What Is It Like to Be a Bass? Red Herrings, Fish Pain and the Study of Animal Sentience
By Vasco Grilo🔸 @ 2026-06-15T16:31 (+19)
This is a linkpost to https://www.frontiersin.org/journals/veterinary-science/articles/10.3389/fvets.2022.788289/full
This is a linkpost for What Is It Like to Be a Bass? Red Herrings, Fish Pain and the Study of Animal Sentience by Georgia Mason and Michelle Lavery, which was originally published in Frontiers in Veterinary Science on 27 April 2022. The abstract and last section are below. The article has a section discussing promising ways of assessing sentience (here is some brief context). Relatedly, I liked the article In search for consciousness in animals: Using working memory and voluntary attention as behavioral indicators by Andreas Nieder, which was published in Neuroscience and Biobehavioral Reviews on 9 September 2022. Thanks to Michael St. Jules for pointing me to both articles.
Abstract
Debates around fishes' ability to feel pain concern sentience: do reactions to tissue damage indicate evaluative consciousness (conscious affect), or mere nociception? Thanks to Braithwaite's research leadership, and concerns that current practices could compromise welfare in countless fish, this issue's importance is beyond dispute. However, nociceptors are merely necessary, not sufficient, for true pain, and many measures held to indicate sentience have the same problem. The question of whether fish feel pain – or indeed anything at all – therefore stimulates sometimes polarized debate. Here, we try to bridge the divide. After reviewing key consciousness concepts, we identify “red herring” measures that should not be used to infer sentience because also present in non-sentient organisms, notably those lacking nervous systems, like plants and protozoa (P); spines disconnected from brains (S); decerebrate mammals and birds (D); and humans in unaware states (U). These “S.P.U.D. subjects” can show approach/withdrawal; react with apparent emotion; change their reactivity with food deprivation or analgesia; discriminate between stimuli; display Pavlovian learning, including some forms of trace conditioning; and even learn simple instrumental responses. Consequently, none of these responses are good indicators of sentience. Potentially more valid are aspects of working memory, operant conditioning, the self-report of state, and forms of higher order cognition. We suggest new experiments on humans to test these hypotheses, as well as modifications to tests for “mental time travel” and self-awareness (e.g., mirror self-recognition) that could allow these to now probe sentience (since currently they reflect perceptual rather than evaluative, affective aspects of consciousness). Because “bullet-proof” neurological and behavioral indicators of sentience are thus still lacking, agnosticism about fish sentience remains widespread. To end, we address how to balance such doubts with welfare protection, discussing concerns raised by key skeptics in this debate. Overall, we celebrate the rigorous evidential standards required by those unconvinced that fish are sentient; laud the compassion and ethical rigor shown by those advocating for welfare protections; and seek to show how precautionary principles still support protecting fish from physical harm.
Discussion and Conclusions: Applying Our Approach To The Fish Pain Debate
What is it like to be a bass? Some claim that fish have rich experiential lives, able to feel pain, fear, and possibly joy [e.g., (12, 13, 24, 184, 185)]. Others claim that fish are essentially unconscious zombies: that being a bass is like nothing, because fish have no phenomenal experience, not even sight [(6); Key in prep., pers. comm.]. The “fish pain” debate is thus rather polarized. Furthermore, discussion and comment still outweigh new data: only 43% of the citers of Sneddon et al. (9) are experimental papers. This research inertia may stem, at least in part, from the untested assumptions of both “sides”. And of course, it also reflects that the whole broad field of P-consciousness is extremely challenging, rife with complexity, debate, and struggles to tackle what is infamously known as “the hard problem”.
As we have reviewed, P-consciousness is currently impossible to measure, and impossible to assess in non-humans. Indeed even in humans, assessment is imperfect, relying on the veracity and accuracy of self-report. This makes the claims at both these extremes too strong – and the high levels of agnosticism about fish abilities to feel pain, appropriate. Perfect, “bullet-proof” diagnostic markers of sentience simply do not exist, at least as yet. Elwood (186) therefore lamented, “the idea of feelings or consciousness … is impossible to access, and [leads to] arguments that cannot be resolved”. We are sympathetic to this, but somewhat more optimistic. We believe one argument that can be resolved is whether sentience matters: it does. Another is whether categorizing fish as sentient will destroy human livelihoods: European data suggest the resulting protections would not necessarily radically hinder animal use (something some may find reassuring, but others, sad). A third argument that we think can be resolved is whether or not to withhold protecting fish until we know they are sentient. The precautionary principle, and the great potential harms done to fish, both indicate that inaction would be ethically worrying; agnosticism about fish sentience is therefore consistent with supporting practical guidelines that choose to protect fish. Indeed, where practices risk extreme pain, we suggest that the guiding question should perhaps not be “is there evidence that this species is sentient?” but instead “are we sure it is not?” Finally, we suggest that empirical research on animal sentience can advance, doing so faster and more constructively, if it is deemed reasonable to treat plants and protozoa, spines disconnected from brains, decerebrate animals and unaware humans (i.e., S.P.U.D. subjects) as not conscious; to treat human self-report as a “gold standard” (as many consciousness researchers do, despite even this being imperfect); and to treat perceptual and evaluative dimensions of P-consciousness as separate and dissociable. We recommend that the questions “Can S.P.U.D subjects do this?” and “In humans, does this always correlate with self-reported feelings, and if so, what type?” are used to screen all potential indicators of animal P-consciousness. Answering these questions will weed out “red herring” measures that fail to distinguish between the sentient and non-sentient, identify types of indicator that best permit strong inference [e.g., sensu (33)], and so assist with both data interpretation and designing new studies.
Applying this approach to current “fish pain” data is revealing: many measures do not survive this screen. For one, S.P.U.D. subjects show diverse unconditioned behavioral responses to noxious stimuli, including avoidance or wound attendance, making it hard to argue that similar responses in fish require awareness or demonstrate true pain rather than mere nociception [cf. e.g., (61, 187), citing (188) on rocking in trout and (189) on tail-beating in zebrafish]. Furthermore, in S.P.U.D. subjects such responses can be modulated, including by analgesics. These means that contrary to several authors [c.f. e.g., (61, 187), citing (188, 190)], the modulation of fish responses to noxious stimuli by analgesics cannot be said to indicate pain over nociception; and the same applies to their modulation by food deprivation [c.f. e.g., (61, 187), citing (191)] or conspecific presence [c.f. e.g., (61, 187), citing (192)]. Furthermore, nor does the conditioning of escape responses to locations where a shock was delivered [c.f. e.g., (61), citing (192)], or of approach responses toward locations where nociceptive input is reduced by analgesics [c.f. e.g., (61, 193), reporting unpublished work on zebrafish], demonstrate pain rather than nociception: S.P.U.D subjects are similarly capable of Pavlovian conditioning [classical/respondant conditioning, for example, dogs starting to salivate after a sound because in the past it was followed by a meal"]. This includes trace conditioning [if the sound stops before the meal shows up] in its broadest sense, often erroneously held up as a marker of P-consciousness [c.f. e.g., (194), citing (195)]: this too occurs in S.P.U.D subjects. Braithwaite and colleagues were therefore right to conclude, in one of Victoria's last papers, “trace conditioning is widespread and by itself does not indicate consciousness” (15), a conclusion echoed in this Special Topic collection by Droege et al. (143). And the same also holds for instrumental learning [operant/instrumental conditioning], where pre-existing innate responses change in timing or form to become more effective (e.g., at avoiding punishment). This means that, for instance, shuttlebox learning by fish, where escape responses become directed to particular locations to avoid shock [e.g., (196)], is also not proof of awareness or pain [context].
Now, authors using such responses to infer pain typically present lists of multiple different responses, to be treated as more convincing if demonstrated en masse [e.g., (197) p. 52; (61, 187, 193); see also (33) “theory neutral” approach]. At first this seems reasonable. However, attributes or responses that are as consistent with a lack of awareness as they are with P-consciousness can have little or no value for inferring true pain, regardless of how numerous they are. Of course, some measures might still be useful if their absence is revealing; in other words if they are deemed necessary for inferring P-consciousness. If a species of fish fails to have nociceptors, for instance [as seems true for at least some elasmobranchs: reviewed by Rose et al. (6), Sneddon (198), Smith and Lewin (199)], then perhaps logically, this is evidence that they cannot feel pain? One challenge would be knowing which these necessary indicators are; and another, distinguishing between true negatives and Type II errors [false negatives]. Nevertheless, thinking more formally about necessary conditions for various forms of animal consciousness (as well as the still elusive sufficient conditions), and parsing these out clearly, would be useful, not least for making assumptions more explicit than they often are.
In contrast, identifying responses that S.P.U.D subjects seem unable to make and ones that in contrast, at least in humans seem to require awareness, highlights other indicators as more useful: better able to permit strong inference (even deduction, if validated using the experiments we suggest). These are measures based on higher order abilities, the self-report of state, working memory, and aspects of operant conditioning. So, what might such indicators reveal about fish?
The apparent intelligence of some fish species [e.g., tool use by tuskfish (200), cooperative hunting by moray eels and groupers (201), numerical competency in angelfish (202), and the mirror self-recognition and episodic memory tasks mentioned above] has sometimes been taken as already sufficient evidence for sentience [e.g., (18)]. For two reasons, we believe this is premature. First, as Massimini and Tononi argue [(45), p. 153], “the mere fact that a behavioral repertoire is complex and “cognitively sophisticated” is not sufficient to clinch the case”. Second, as we have reviewed here, the perceptual and evaluative dimensions of P-consciousness should be considered distinct [following (36)], and these tasks typically provide better evidence of the former than the latter. Nevertheless, the apparent intelligence of such species does bode well for tasks specifically designed to probe sentience, making this an exciting area for future work. Thus, in the section What Could Be Evidence of P-Consciousness and, More Narrowly, Sentience?, we suggested modifying existing tests for self-awareness and episodic memory, in order to make them sensitive to sentience (not just the visual, proprioceptive and temporal P-consciousness that they rely on in their current forms). We hope our outlines for a “mirror test with biting parasite”, and for a “what, where, and in which state?” episodic memory task, indicate how such paradigms could be tweaked – both in new assumption-testing experiments on humans, and in new experiments with animals. In that same section, we also suggested ways in which self-report paradigms could be developed for animals, including fish (since when animals are trained to use their internal states as discriminative stimuli, highly revealing insights can emerge, as the drug discrimination literature reveals). We believe there are ways to avoid such tasks' risks of just capturing blindsight-like guessing with no awareness [see blindsight], and again, that the intelligence of some fish bodes well for applying these. Were such affect-sensitive higher-order tasks applied to fish (especially once better validated), we would therefore recommend starting with these species with impressive cognitive abilities.
What about other, seemingly less impressive cognitive abilities? We and others [e.g., (116)] have suggested operant learning as a relevant topic to explore. Our own suggestions about operating learning arise partly from looking for tasks that S.P.U.D. subjects seem unable to do. Of course, absence of evidence is not evidence of absence: the lack of data from S.P.U.D. subjects on the learning of arbitrary operants could reflect a lack of research effort, not a failure of such attempts to work. This therefore identifies another topic for future research, using both S.P.U.D. subjects and humans whose awareness of reinforcers is manipulated: the assessment of whether operant behavior relies on reinforcers producing conscious affective states. (Though note that for such work, we suggest using plants and temporarily unaware humans over highly invasive decerebrate or spinally-transected animals). If this hypothesis was supported, then at least some fish have already shown that they are sentient. These include Siamese fighting fish trained to swim through hoops to display to rivals [e.g., (203)], barramundi (Lates calcarifer) trained to touch arbitrary targets (204) and goldfish trained to bump a lever for food (205), as well as all farmed species that use demand feeders to deliver pellets.
More stringently, it could be that not all operant learning requires sentience [it demonstrably does not require consciousness of the rewards and punishments], but that only some sub-types do. Thus, it could be that Ginsburg and Jablonka's (37) hypothesis about second-order conditioning [context] is correct: again something not yet known, but amenable to empirical test. If their hypothesis is supported [it may not be], then again there are already some cases demonstrating conditioned responses to secondary reinforcers by fish. In one, for example, Foerder (204) successfully used shaping to train a barramundi to swim 10 feet to touch a secondary reinforcer (a target previously paired with food). In another, an extraordinary experiment demonstrated the abilities of goldfish to learn to jump over a hurdle to modify a CS [conditioned stimulus, like a sound that makes dogs salivate] (lights that must either match or not), in order for this CS to predict no shock rather than shock (206). Thus, if future research does show that second-order conditioning relies on the conscious awareness of reinforcers, then these fish are displaying good evidence for positive affect and pain, respectively. What about the other sub-type of operant learning that we highlighted: goal-directed? It is not yet known whether goal-directed operant responding for food, say, require this reward to induce consciously-experienced positive states, but again this idea is testable. And were the hypothesis supported, then species with flexible forms of foraging, like those using the cooperation and tool use mentioned above, seem ideal for formally investigating whether they can show goal-directed operant behavior, in novel experiments specifically designed to assess this ability.
Operant learning is just one of the many topics tackled in considerable ongoing research by human consciousness researchers (with the use of discriminative stimuli, and working memory being others). We recommend following this fascinating work, for example by attending meetings such those hosted by the Association for the Scientific Study of Consciousness: learning about this complex, fast-moving human research is both useful and humbling. Furthermore, increased contact with and interest from those working on animal welfare might encourage such researchers to focus more on understanding sentience (since currently, most of their research efforts focus on perceptual forms of P-consciousness). We hope that our suggestions represent a constructive contribution to the deeply interesting, important, but sometimes frustrating field of animal consciousness. Inspired by Victoria, one of the first to wade into this fray, we look forward to the refinement of these ideas and improved methodologies for probing the puzzle of animal sentience.
VeryJerry @ 2026-06-17T17:58 (+3)
Based on everything you've looked into, what odds do you put on fish sentience?
I would add, I'm not convinced that all "unconscious" states in humans are equal. I've heard reports of people who "wake up" during surgery, but were still unable to move. Note that I haven't looked into this in any depth and it may very well be an urban legend. It could be that some anesthetics knock out ability to move and long term memory, such that it appears as though the person is not feeling pain and they can't report pain they don't remember, although they may have felt it at the time.
Vasco Grilo🔸 @ 2026-06-17T18:27 (+3)
Hello.
Based on everything you've looked into, what odds do you put on fish sentience?
I speculate carp have around 50 % (= (0.70 + 0.40)/2) chance of being sentient, but I have little reason to expect my intuitions are calibrated. I feel like anything from 0.1 % to 90 % is reasonable. In any case, I can see the welfare of fish being very close to 0 even if they are sentient. So I would rather prioritise decreasing uncertainty about their sentience and intensity of their experiences over investing in interventions helping fish.
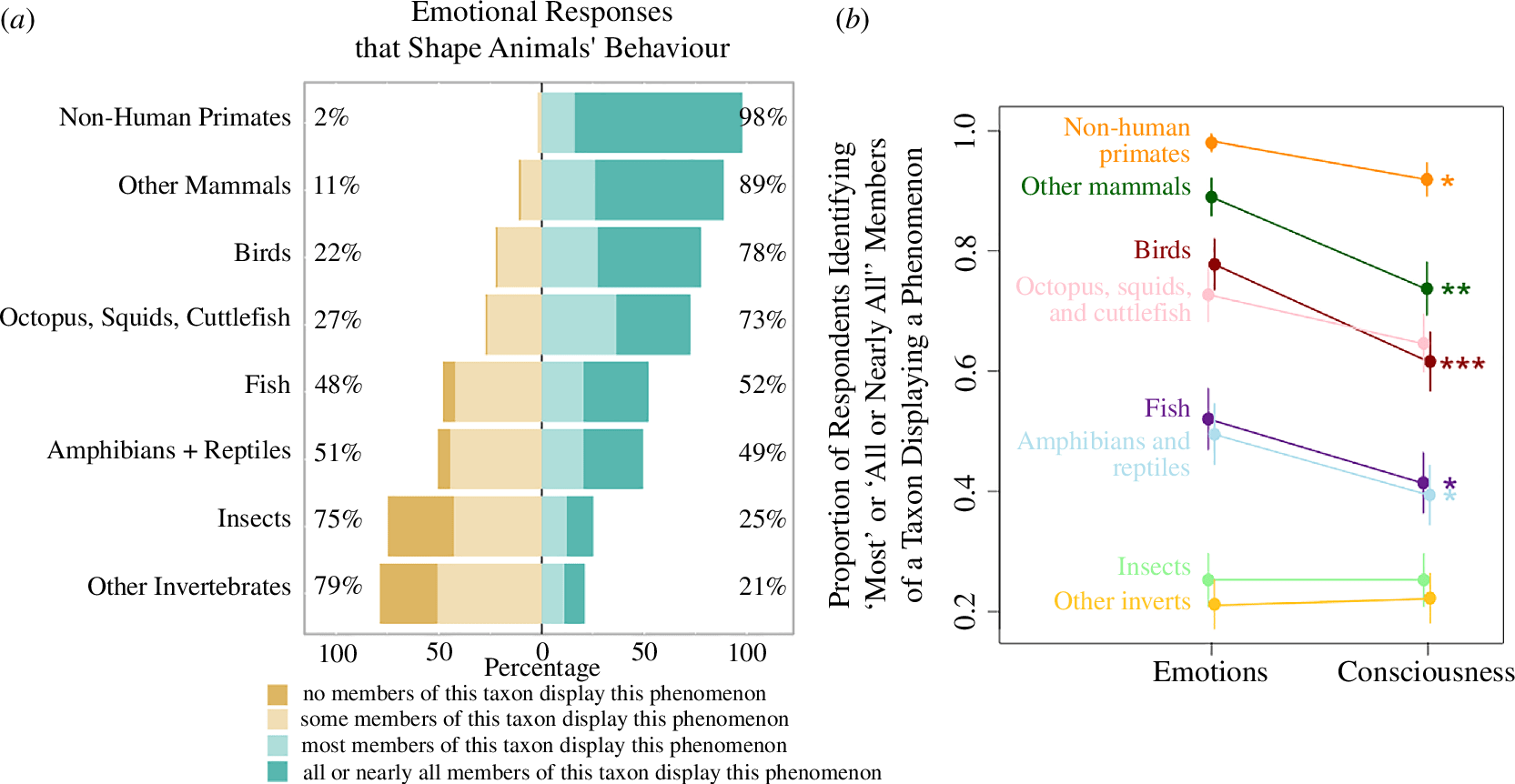
Here is some additional context you may be interested in about the likelihood of fish being sentient. Bob Fischer's book about comparing welfare across species presents an estimate of 70 % for the sentience of carp. Among the people surveyed in Zipple et al. (2024), as illustrated in Figure 1 below, around 40 % guessed "most" or "all or nearly all" fish are conscious. Importantly, "we defined consciousness for respondents, using a ‘self-consciousness’ or ‘self-awareness’ definition (an animal being aware of its own existence [33–35])".
I would add, I'm not convinced that all "unconscious" states in humans are equal. I've heard reports of people who "wake up" during surgery, but were still unable to move.
Anesthesia awareness is a real (though rare) phenomenon. However, note that people who are awake would no longer qualify as unconscious under the definition of the article.
NickLaing @ 2026-06-17T18:41 (+5)
I appreciate this paragraph, especially the extreme uncertainty and the welfare of fish being potentially close to zero even if sentient.
"I speculate carp have around 50 % (= (0.70 + 0.40)/2) chance of being sentient, but I have little reason to expect my intuitions are calibrated. I feel like anything from 0.1 % to 90 % is reasonable. In any case, I can see the welfare of fish being very close to 0 even if they are sentient. So I would rather prioritise decreasing uncertainty about their sentience and intensity of their experiences over investing in interventions helping fish."
What kind of research do you think could decrease uncertainty about their sentience? I'm assuming you mean behavioural research? I think this kind of research is helpful to reduce uncertainty but doesn't necessarily address fundamental questions about how brains work.
Vasco Grilo🔸 @ 2026-06-17T20:40 (+12)
Hi Nick.
What kind of research do you think could decrease uncertainty about their sentience?
The kind of research described in this section of the article. Tasks involving working memory, aspects of operant conditioning, self-report of sensation, or higher order consciousness (like the “mirror test with biting parasite”, which offers stronger evidence for sentience than the standard mirror test). Below is some context from the article about why these tasks are promising. I bolded the 4 types of tasks.
here we suggest four types of candidate measure that seem particularly promising as indicators of sentience. One concerns working memory, and another, forms of operant conditioning: two types of ability that S.P.U.D. subjects seem not to convincingly have. A third type of candidate measure concerns self report. As we saw in the section Red Herrings: Responses That Do Not Require Sentience, unaware humans, and also monkeys with blindsight, report sensing nothing (thence having no P-consciousness), even when presented with stimuli that elicit other kinds of response – with the human subjects consistently describing themselves as merely guessing. Finally, higher order cognitive processes such as episodic memory and self-recognition, can yield insights into animal awareness. These have not been studied in S.P.U.D. subjects, and this would likely be impossible, but are arguably reliant on perceptual consciousness. We review each of these below, and also outline how they could be modified to now address questions about sentience: the ethically relevant dimension of P-consciousness at the heart of the fish pain debate. We also suggest how their validity as indicators of sentience could be assessed.
In addition, tasks involving endogenous/voluntary attention, like Posner’s spatial cueing task, as discussed in Nieder (2022).
I'm assuming you mean behavioural research?
All of the above tasks look into behaviour. However, passing them would be stronger evidence for sentience than passing tasks that have been passed by SPUDs (spines disconnected from brains, humans in unaware states, decerebrate mammals and birds, or organisms lacking a nervous system like plants and protozoa).
I think this kind of research is helpful to reduce uncertainty but doesn't necessarily address fundamental questions about how brains work.
Note that research looking for the neural correlates of consciosness (NCCs) in animals relies on studying the behaviour of humans. In particular, to understand which states of the human brain are responsible for consciousness. Nieder (2022) says there is no evidence for the neural correlates of consciousness in animals besides mammals and birds.

NickLaing @ 2026-06-18T05:07 (+5)
My apologies @Vasco Grilo🔸 I was lazy there and didn't read the whole article properly. Thanks for pointing out the important areas. Working memory and operant conditioning don't carry any weight with me personally, I struggle to connect those with a meaningfully increased chance of pain experience. For example the portia spider has a very advanced working memory for very specific tasks which makes sense evolutionary, as do many other organisms, but i can't imagine why these specific evolutionary advantages would also lead to sentience?
"Finally, higher order cognitive processes such as episodic memory and self-recognition, can yield insights into animal awareness. These have not been studied in S.P.U.D. subjects, and this would likely be impossible"
These higher order processes would be most convincing for me than the memory stuff, especially deeper levels of self recognition which I feel like could maybe be tested?
Perhaps a silly question, but I thought the mirror/parasite mirror test was related to "self recognition", but you seem to mention that as something different.
Vasco Grilo🔸 @ 2026-06-18T08:52 (+3)
My apologies @Vasco Grilo🔸 I was lazy there and didn't read the whole article properly. Thanks for pointing out the important areas.
No worries. I could have highlighted them more too. So thanks for asking. I linked to the relevant section in this comment, but I have now linked to it at the top of the post too. "The article has a section discussing promising ways of assessing sentience (here is some brief context [my comment just above])".
Nitpick. I get email notifications of comments on my posts, and replies to my comments even if I am not tagged. I think this is the default setting on the EA Forum. If I am tagged in those cases, I get 2 email notifications, although this does not bother me.
Working memory and operant conditioning don't carry any weight with me personally, I struggle to connect those with a meaningfully increased chance of pain experience.
I would also give them the least weight among the 4 indicators, but I am not confident they should have negligible weight. I think negligible weight makes more sense for classical/respondant conditioning, as SPUDs are capable of this.
In addition, I believe some types of working memory and operant conditioning should have more weight than others. Here is some brief context from the post.
More stringently, it could be that not all operant learning requires sentience [it demonstrably does not require consciousness of the rewards and punishments], but that only some sub-types do. Thus, it could be that Ginsburg and Jablonka's (37) hypothesis about second-order conditioning [context] is correct: again something not yet known, but amenable to empirical test. If their hypothesis is supported [it may not be], then again there are already some cases demonstrating conditioned responses to secondary reinforcers by fish.
These higher order processes would be most convincing for me than the memory stuff, especially deeper levels of self recognition which I feel like could maybe be tested?
Perhaps a silly question, but I thought the mirror/parasite mirror test was related to "self recognition", but you seem to mention that as something different.
Both the mirror test and “mirror test with biting parasite” test self-recognition, but the latter offers stronger evidence for sentience. I asked Gemini 3.5 Flash to contrast the 2 tests with respect to this given the article's description of the “mirror test with biting parasite”. The answer is below. I think it is good, except that it is overconfident (as it is often the case with LLMs). Words like "proves" and "demonstrates" should be replaced with "offers stronger evidence (relative to the standard mirror test) for".
The standard mirror test is widely used to check for self-awareness, but it doesn't tell us if an animal can actually feel things like pleasure or pain.
Mason and Lavery (2022) proposed a modified version to bridge this gap. Here is how the two tasks contrast and why the new one offers stronger evidence for sentience.
The Two Tasks Contrasted
- The Standard Mirror Test (Affectively Neutral): An animal is marked with a spot (like a dot of odorless, painless paint) in a place it can only see using a mirror (like its forehead). If the animal looks in the mirror and then grooms, touches, or scrapes its own body where the mark is, it passes. This proves the animal understands that the reflection is "me" rather than another animal, showing a baseline of physical and visual self-awareness. However, because the mark doesn't feel like anything, the task is affectively neutral—it doesn't involve emotions or feelings.
- The "Mirror Test with a Biting Parasite" (Affectively Charged): This version introduces feelings. Instead of a neutral dot, the animal is trained to associate different visual marks with physical comfort or discomfort. For example:
- The animal is exposed to a black triangle mark that is physically uncomfortable or painful (simulating a biting parasite).
- It is also exposed to a white circle mark that causes no discomfort at all.
- Finally, the researchers place these marks on the animal's body without any actual pain or discomfort attached, in a spot the animal can see directly or via a mirror. The test checks if the animal reacts with greater urgency, anxiety, or scraping behaviors toward the "nasty" black triangle than the "benign" white circle, purely based on what it looks like.
Why the Parasite Test Provides Stronger Evidence for Sentience
To understand why the parasite version is stronger, it helps to separate two concepts: awareness and sentience.
Passing the standard mirror test proves awareness—specifically, the cognitive ability to map visual reflections to one's own physical body. But critics argue that a highly complex, non-sentient robot or AI could theoretically pass this test using pure geometry and visual processing, without experiencing any inner feelings.
The biting parasite test targets phenomenal consciousness (P-consciousness) and affective states—the actual capacity to subjectively feel pain, fear, or distress.
By showing that an animal doesn't just recognize a mark, but specifically anticipates and reacts to the remembered emotional and painful quality of that mark, the test demonstrates that the animal experiences the world through a lens of "good" and "bad" feelings. It links visual self-recognition directly to an inner emotional state (the desire to avoid pain), providing much sturdier proof that the animal is truly sentient and capable of suffering.
Vasco Grilo🔸 @ 2026-06-16T07:55 (+2)
I think impact-focussed funders underrate research on animal sentience, and, more broadly, on comparing welfare across species. I believe there is huge uncertainty, and ways of decreasing it. Here is some context about my uncertainty. In Bob Fischer’s book about comparing welfare across species, the tentative sentience-adjusted welfare range of shrimps is 8.0 % of that of humans. Welfare range is defined there as the difference between the maximum and minimum welfare per unit time among “realistic biological possibilities”. For sentience-adjusted welfare ranges proportional to “individual number of neurons”^“exponent”, and “exponent” from 0 to 2, which covers the best guesses that I consider reasonable, the sentience-adjusted welfare range of shrimps is 10^-12 to 1 times that of humans.