Cost-effectiveness accounting for soil nematodes, mites, and springtails
By Vasco Grilo🔸 @ 2025-06-03T16:43 (+78)
Side note. I am looking for work, and welcome suggestions for posts.
Summary
- I think the impact of the vast majority of interventions is driven by effects on wild animals. In particular, by effects from land use change on soil nematodes, mites, and springtails, which are the most abundant terrestrial animals. There are 4.4*10^20 top soil nematodes, and 10^19 soil arthropods, “≈95% of which are soil mites and springtails”.
- I estimate random soil nematodes, mites, and springtails have (expected) welfare ranges (as fractions of that of humans) of 6.68*10^-6, 3.65*10^-5, and 6.29*10^-5, which are 0.334 %, 1.83 %, and 3.15 % of Rethink Priorities’ (RP’s) mainline welfare range of silkworms.
- I calculate soil nematodes, mites, and springtails have (in expectation) a welfare of -4.48*10^-6, -1.61*10^-5, and -2.39*10^-5 QALY/animal-year, and an annual welfare of -306 k, -14.2 k, and -10.6 k times that of humans. The annual welfare of soil nematodes being 12.8 times that of soil arthropods suggests the former have been unfairly dismissed relative to the latter by people who care a lot about wild animal welfare.
- Crops are the biome with the 3rd highest welfare among the 10 I analysed. I infer increasing cropland only decreases welfare if it replaces pasture, or deserts, and xeric shrublands, which are the 2 biomes with a lower density of soil animals than crops.
- My results imply cropland replacing tropical and subtropical grasslands, savannas, and shrublands decreases the living time of soil animals by 1.73 M animal-years per m2-year, and increases their welfare by 8.15 QALY/m2-year, of which 93.2 % comes from increasing the welfare of soil nematodes. I multiply the increase in QALY/m2-year by changes in cropland in m2-year/$ to estimate the cost-effectiveness accounting only for soil animals. I get the following cost-effectiveness accounting for target beneficiaries and soil animals as a fraction of the past cost-effectiveness of Shrimp Welfare Project’s (SWP’s) Humane Slaughter Initiative (HSI):
- For donating to the High Impact Philanthropy Fund (HIPF) from the Centre for Exploratory Altruism Research (CEARCH), 20.9 (13.4 kQALY/$).
- For GiveWell’s top charities, 1.74 (1.11 kQALY/$).
- For Broiler welfare corporate campaigns, 1.20 (767 QALY/$).
- For buying beef, 65.8 % (420 QALY/$).
- For cage-free corporate campaigns, 21.6 % (138 QALY/$).
- For Veganuary in 2024, -31.8 % (-203 QALY/$).
- For School Plates in 2023, -7.79 (-4.97 kQALY/$).
- I am uncertain about whether each of the above is beneficial or harmful. Likewise for other interventions aiming to help vertebrates which change cropland. I estimate their effects on the target beneficiaries are negligible compared with those on soil animals, and it is unclear whether these have positive or negative lives, which results in cost-effectiveness distributions with positive and negative heavy tails. I calculate soil nematodes, mites, and springtails have negative lives with a probability of 58.7 %, 55.8 %, and 55.0 %.
- I consider it very worth it to decrease the uncertainty about how interventions affect the living time of soil nematodes, mites, and springtails, and about their welfare in QALY/animal-year. In particular, it is crucial to know whether they have positive or negative lives. I am not aware of any organisations working reasonably directly on this. I wonder how much money RP or Wild Animal Initiative (WAI) would need to make some progress with targeted projects.
- I suspect donating to HIPF increases welfare more cost-effectively than to the organisations working on invertebrate welfare I had recommended, although I believe these are much less likely to be harmful. I estimate donating to HIPF decreases the living time of soil nematodes, mites, and springtails by 2.84 billion animal-years per $.
- I no longer believe animal welfare should be more prominently promoted by effective giving initiatives (EGIs). Increasing donations to interventions whose target beneficiaries are humans is easier, resulting in more additional donations per $ spent, and I estimate the most cost-effective ones are roughly as cost-effective as HSI has been.
- I do not expect the meat-eating problem to be problematic due to positive effects on wild animals.
- As I expected, my analysis indicates the impact of chicken welfare reforms is driven by effects on wild animals. I estimate broiler welfare and cage-free corporate campaigns benefit soil animals 458 and 29.0 times as much as they benefit chickens. I think the same applies to any intervention targeting vertebrates which changes the consumption of feed or food, especially if it mainly aims to increase/decrease positive/negative vertebrate-years.
- My best guess is that decreasing the consumption of animal-based foods is harmful. I estimate School Plates in 2023, and Veganuary in 2024 harmed soil animals 5.59 k and 3.69 k times as much as they benefited farmed animals.
- Donating more and better becomes less valuable than I had suggested after accounting for effects on soil animals.
- Effects on soil animals cannot be neglected just because they are uncertain.
Introduction
I have been assuming interventions aiming to increase or decrease the number of wild animals are not worth it because I am uncertain about whether they have positive or negative lives. However, my analysis illustrating chicken welfare reforms have larger effects on wild arthropods than chickens got me thinking, and making a few Fermi estimates suggesting they could be close to as cost-effective as SWP’s HSI, which is saying a lot. SWP is the only organisation I recommended in the analysis of the chicken welfare reforms not focussing overwhelmingly on research.
Here is one of the Fermi estimates I did in a piece of paper, which is why I used rounded numbers. A change from temperate forests to crops of 300 m2-year per beef-kg, cost of 10 $ per beef-kg, change in population of 100 k arthropod-year/m2-year (= (100 - 20)*10^3) more mites in temperate forests than crops from Table S4 of Rosenberg et al. (2023), and increase in welfare of 5.00*10^-5 QALY/arthropod-year imply a cost-effectiveness of 150 QALY/$ (= 300/10*100*10^3*5.00*10^-5), 1/4 (= 150/639) of my estimate for HSI.
Uncertainty about whether wild arthropods have positive or negative lives implies a welfare close to 0, but exactly how close matters. Refusing to quantify is refusing to think (about trade-offs), and there are interventions that achieve huge changes in arthropod-years per $.
I think the impact of the vast majority of interventions is driven by effects on wild animals. In particular, by effects from land use change on soil nematodes, mites, and springtails, which are the most abundant terrestrial animals. There are 4.4*10^20 top soil nematodes, and 10^19 soil arthropods, “≈95% of which are soil mites and springtails”. van den Hoogen et al. (2019) got the number of soil nematodes adding estimates by pixel, a very small area, obtained from environmental variables. They “focus on the top 15 cm of soil, which is the most biologically active zone of soils”, so I asked the 1st 2 authors about the total number of soil nematodes. Stefan Geisen, the 2nd author, clarified they focussed on the most active layer, which was sometimes more or less than 15 cm, and guessed their estimate accounts for 90 % of all soil nematodes. I divided their estimate by 90 % to estimate 4.89*10^20 soil nematodes. Rosenberg et al. (2023) got the number of soil arthropods adding estimates by biome obtained from multiplying their area by the mean density across their sites.
In this post, I estimate the welfare range and welfare of random soil nematodes, mites, and springtails, and the cost-effectiveness of some interventions accounting for target beneficiaries and soil animals. Here are my calculations.
I have not considered the effects on marine nematodes and arthropods, which are super abundant too. From Table S1 of Bar-on et al. (2018), there are 10^21 nematodes (2.05 times my number for soil nematodes), and 10^20 marine arthropods (10.0 times my number for soil arthropods). Nevertheless, I guess the effects on marine nematodes and arthropods, as well as on all potential sentient beings on Earth considered together, are beneficial/harmful if the direct effects from land use change on soil animals are beneficial/harmful, such that including other considerations would not revert my conclusions. I also tend to agree with Bentham’s Bulldog that increasing human-years benefits wild animals longterm, although I expect the effects over the 1st 100 years (after the spending) to cover the vast majority of the total impact.
Welfare ranges
I have been relying on RP’s mainline welfare ranges to estimate the welfare of animals. RP has not produced estimates for soil nematodes, mites, or springtails, which forces me to improvise. I get welfare ranges for the modal (most common) soil nematode, mite, and springtail based on the relationship between RP’s mainline welfare ranges, and the number of neurons of the analysed species. As part of RP’s moral weight project, which produced their mainline welfare ranges, Adam Shriver wrote a report arguing the number of neurons should not be used as a proxy for moral weight. The post summarising says “there is no straightforward empirical evidence or compelling conceptual arguments indicating that relative differences in neuron counts within or between species reliably predicts welfare relevant functional capacities”. Yet, it turns out non-linear functions of the number of neurons predict RP’s mainline welfare ranges fairly well.
I consider 2 different models. Denoting the welfare range by WR, and the number of neurons by N:
- WR = a*N^b, which is equivalent to ln(WR) = ln(a) + b*ln(N). I determine a and b with a linear regression of the logarithm of the welfare range on the logarithm of the number of neurons, which has:
- A coefficient of determination (R^2) of 62.3 %.
- A slope of 0.308, which implies the welfare range is proportional to N^0.308.
- WR = ln(1 + c*N), which is equivalent to e^WR - 1 = c*N. I determine c with a linear regression with null intercept of the exponential of the welfare range minus 1 on the number of neurons, which has:
- An R^2 of 81.7 %.
- A slope of 2.00*10^-11, which implies the welfare range is proportional to ln(1 + 2.00*10^-11*N).
Here is the representation of the 1st regression. I used RP’s numbers of neurons, but I suspect that of silkworms is too high. It is 95.6 % (= 860*10^3/(900*10^3)) of that of bees, and I guess these have much more neurons than silkworms.
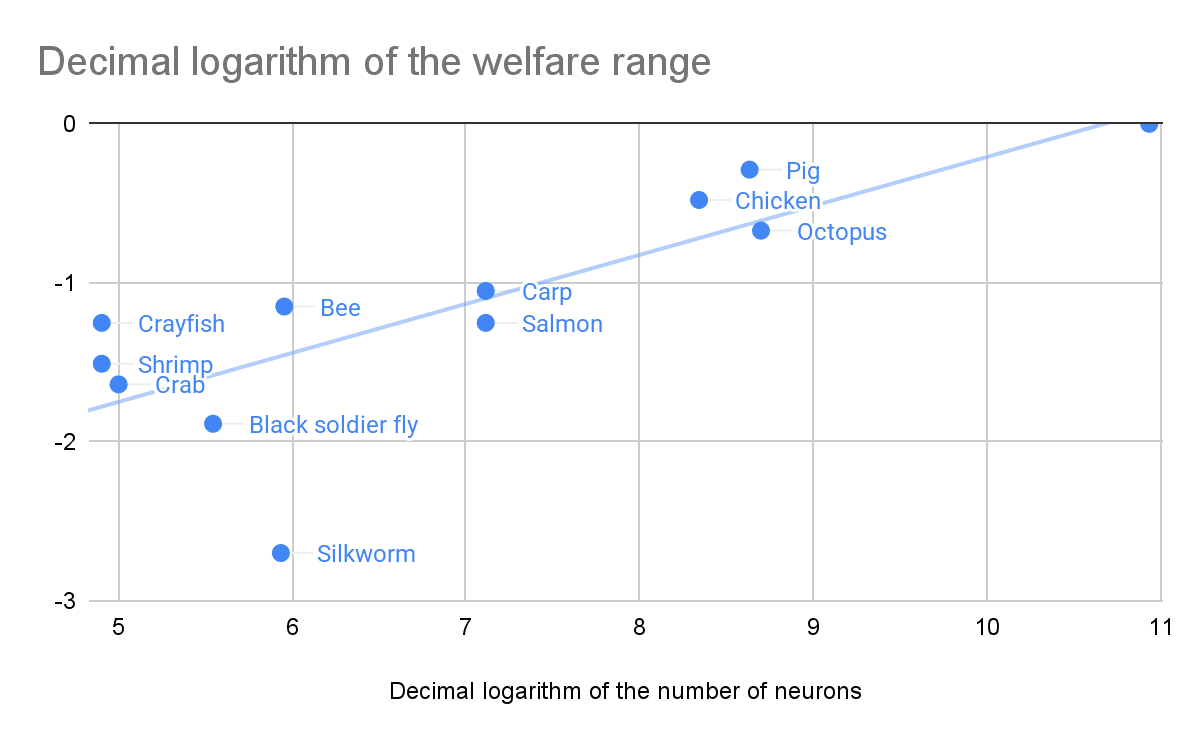
I set up the models such that the welfare range is 0 for no neurons. In addition, I think the welfare ranges of the modal soil nematode, mite, and springtail should be lower than RP’s lowest mainline welfare ranges of silkworms. So I scale this to estimate those ranges:
- WR = “RP’s mainline WR of silkworms”*(N/“N of silkworms”)^b (method 1).
- WR = “RP’s mainline WR of silkworms”*ln(1 + c*N)/ln(1 + c*“N of silkworms”) (method 2).
I combine the results of the 2 models with a geometric mean weighted by R^2. Aggregating lognormal distributions whose logarithms have the same standard deviation with the continuous version of the geometric mean of odds results in a distribution whose mean is equal to the geometric mean of the means of the lognormal distributions.
I asked Gemini 2.5 Pro (preview) on 12 May 2025 about the modal soil nematode, mite, and springtail, and the mean of Gemini’s best guess distribution for their number of neurons. The results are below. “E” stands for “*10^”.
| Animal | Species (life stage) | Number of neurons | Number of neurons as a fraction of that of silkworms | Welfare range (method 1) | Welfare range (method 2) | Welfare range | Welfare range as a fraction of that of silkworms |
| Modal soil nematode | Caenorhabditis elegans (L1 juvenile) | 240 | 0.0279% | 1.50E-04 | 5.58E-07 | 6.68E-06 | 0.334% |
| Modal soil mite | Tectocepheus velatus (larva) | 2.75E+03 | 0.320% | 3.24E-04 | 6.40E-06 | 3.65E-05 | 1.83% |
| Modal soil springtail | Folsomia candida (1st instar) | 6.00E+03 | 0.698% | 4.15E-04 | 1.40E-05 | 6.29E-05 | 3.15% |
| Random soil arthropod | Not defined | 3.83E+03 | 0.446% | 3.55E-04 | 8.91E-06 | 4.53E-05 | 2.27% |
The estimates above respect the earliest life stage excluding eggs, which have less neurons than later stages. My estimates for a random soil arthropod assume 2/3 of soil arthropods are modal mites, and 1/3 are modal springtails. According to Rosenberg et al. (2023), soil mites and springtails account for 95 % of soil arthropods, soil mites account for 2/3 of soil mites and springtails, and soil springtails for 1/3.
I assume the welfare range of random soil nematodes, mites, and springtails is very similar to that of modal soil nematodes, mites, and springtails, as simpler animals tend to be more abundant. So I estimate random soil nematodes, mites, and springtails have welfare ranges matching the ones in the table above of 6.68*10^-6, 3.65*10^-5, and 6.29*10^-5, which are 0.334 %, 1.83 %, and 3.15 % of RP’s mainline welfare ranges of silkworms.
Welfare per animal-year, and annual welfare
I asked Gemini 2.5 Pro (preview) on 14 May 2025 about the welfare per animal-year of random wild animals of the aforementioned species and life stages as a fraction of that of fully healthy animals of the same species and life stage. Here is the prompt I used.
Hi Gem,
What are your best guesses for:
- The hedonistic welfare per animal-year of random wild Caenorhabditis elegans L1 juveniles as a fraction of that of fully healthy Caenorhabditis elegans L1 juveniles whose mortality is negligible.- The hedonistic welfare per animal-year of random wild Tectocepheus velatus larvae as a fraction of that of fully healthy Tectocepheus velatus larvae whose mortality is negligible.
- The hedonistic welfare per animal-year of random wild Folsomia candida 1st instars as a fraction of that of fully healthy Folsomia candida 1st instars whose mortality is negligible.
Please provide point estimates representing the means of your best guess distributions. Positive/Negative estimates imply lives with more happiness/suffering than suffering/happiness, such that the animals wanting to maximise their own welfare would prefer existing over not existing.
I have obtained the following estimates for farmed chickens (https://forum.effectivealtruism.org/posts/8FqWSqv9AeLowgajn/cost-effectiveness-of-corporate-campaigns-for-chicken#Welfare_per_living_time):
- For broilers in conventional (fast-growth) and reformed (slow-growth; Better Chicken Commitment) scenarios, -2.27 and -0.161.
- For laying hens in conventional cages and cage-free aviaries, -1.69 and -0.333.
In addition, I estimate a value of 0.885 for humans in 2021, 1 minus the 0.115 years lost due to disability (YLD) [described here] per capita in 2021 [from the Global Burden of Disease Study (GBD)].
Gemini provided best guesses for soil nematodes, mites, and springtails of -67 %, -44 %, and -38 %, which are 1.60, 1.05, and 0.905 times Ambitious Impact’s estimate of -42 % for wild bugs based on their deprecated welfare points system. My sense is also that most people working on wild animal welfare would guess soil nematodes, mites, and springtails have negative lives. I defer to Gemini’s estimates implying modal soil nematodes, mites, and springtails have negative lives, but there is lots of uncertainty. My very tentative best guess distributions are normal ones with Gemini’s means, and 5th and 95th percentiles equal to the means minus and plus 5. These distributions imply probabilities of negative welfare slightly above 50 %, as shown in the table below. I would not be surprised if soil nematodes, mites, and springtails had positive lives.
| Animals | Welfare per animal-year as a fraction of that of fully healthy animals | |||||||
| Mean | Mean as a fraction of Ambitious Impact's estimate for wild bugs | Difference between the 5th percentile and mean | Difference between the 95th percentile and mean | 5th percentile | 95th percentile | Standard deviation | Probability of being negative | |
| Soil nematodes | -67.0% | 1.60 | -5.00 | 5.00 | -5.67 | 4.33 | 3.04 | 58.7% |
| Soil mites | -44.0% | 1.05 | -5.00 | 5.00 | -5.44 | 4.56 | 3.04 | 55.8% |
| Soil springtails | -38.0% | 0.905 | -5.00 | 5.00 | -5.38 | 4.62 | 3.04 | 55.0% |
I had initially planned to estimate the means above based on guesses for the intensity of and time in the categories of pain and pleasure defined by the Welfare Footprint Institute (WFI), similarly to what I have done before. I tried to obtain estimates for the time using WFI’s GPT Hedonic Track (HT), but they did not make sense. HT said 100 % of Caenorhabditis elegans die during the L1 stage without seemingly being aware that exactly 100 % dying would imply the extinction of the species. Moreover, HT estimated the modal Caenorhabditis elegans surviving L1 experiences less annoying pain than the modal individual of that species not surviving that stage, and as much hurtful, disabling, and excruciating pain, joy, euphoria, and bliss, whereas I guess survivors should experience at least more disabling and excruciating pain due to avoiding death.
I compute the welfare in QALYs per animal-year multiplying the welfare range by the welfare per animal-year as a fraction of that of fully healthy animals. I determine the welfare in QALYs per year multiplying the population by the QALYs per animal-year. The results are below, including for humans and soil animals. I suppose these are just soil nematodes and arthropods, which is practically true.
| Animals | Population | Welfare range | Welfare per animal-year as a fraction of that of fully healthy animals | QALYs per animal-year | QALYs per year | QALYs per year as a fraction of those of humans |
| Humans | 8.09E+09 | 1.00 | 88.5% | 0.885 | 7.16E+09 | 100% |
| Soil nematodes | 4.89E+20 | 6.68E-06 | -67.0% | -4.48E-06 | -2.19E+15 | -3.06E+05 |
| Soil mites | 6.33E+18 | 3.65E-05 | -44.0% | -1.61E-05 | -1.02E+14 | -1.42E+04 |
| Soil springtails | 3.17E+18 | 6.29E-05 | -38.0% | -2.39E-05 | -7.57E+13 | -1.06E+04 |
| Soil arthropods | 1.00E+19 | 4.31E-05 | -39.9% | -1.72E-05 | -1.72E+14 | -2.40E+04 |
| Soil animals | 4.99E+20 | 7.41E-06 | -66.5% | -4.92E-06 | -2.46E+15 | -3.43E+05 |
I calculate soil nematodes, mites, and springtails have a welfare of -4.48*10^-6, -1.61*10^-5, and -2.39*10^-5 QALY/animal-year, and an annual welfare of -306 k, -14.2 k, and -10.6 k times that of humans. The annual welfare of soil nematodes being 12.8 (= -306*10^3/(-24.0*10^3)) times that of soil arthropods suggests the former have been unfairly dismissed relative to the latter by people who care a lot about wild animal welfare.
Welfare per area
I determine the QALYs per m2-year multiplying the QALYs per animal-year by the number of individuals per m2. For the density of mites and springtails by biome, I use values from Table S4 of Rosenberg et al. (2023), which are represented in Figure 2B below.
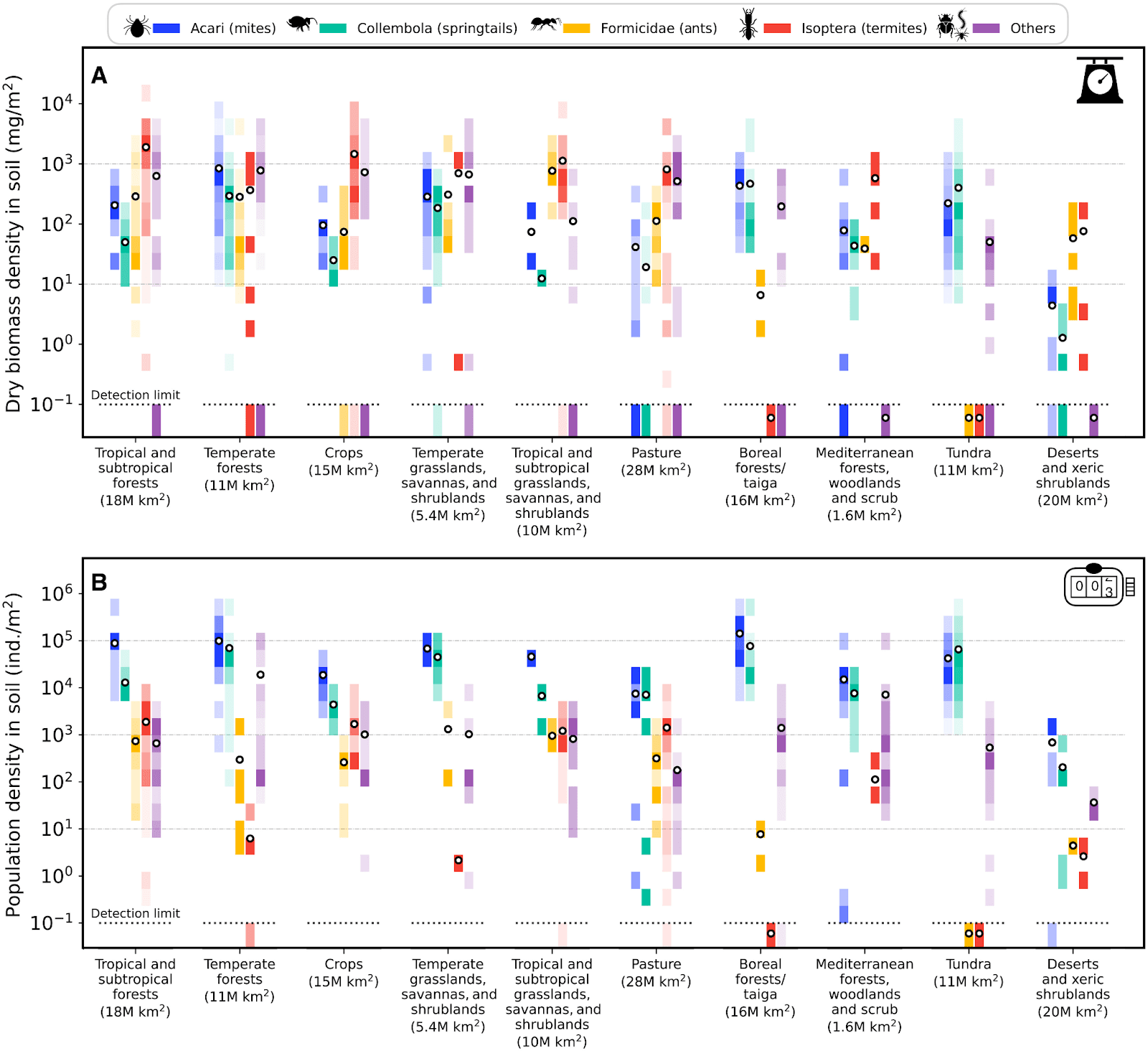
I set the density of nematodes to the product between the sum of the densities of mites and springtails, and 51.5, which is my estimate for the number of soil nematodes as a fraction of the number of soil mites and springtails. Extended Data Figure 4 of van den Hoogen et al. (2019) has a map with the density of nematodes, but the density by biome is only provided in Figure 1b in terms of nematodes per 100 g of dry soil. I asked the corresponding authors of van den Hoogen et al. (2019) and Rosenberg et al. (2023) about a better way of quickly estimating the density of soil nematodes in the biomes of Table S4 of Rosenberg et al. (2023) based on the density of mites and springtails reported there, but the respective 1st authors were not aware of any.
The results are in the table below.
| Biome | Soil nematodes per m² | Soil mites per m² | Soil springtails per m² | Soil arthropods per m² | Soil animals per m² | Decrease in soil animals per m² from replacing the biome with crops | Welfare of soil nematodes (QALY/m²-year) | Welfare of soil mites (QALY/m²-year) | Welfare of soil springtails (QALY/m²-year) | Welfare of soil arthropods (QALY/m²-year) |
| Boreal forests / Taiga | 9.26E+06 | 1.00E+05 | 8.00E+04 | 1.80E+05 | 9.44E+06 | 8.18E+06 | -41.5 | -1.61 | -1.91 | -3.52 |
| Crops | 1.24E+06 | 2.00E+04 | 4.00E+03 | 2.40E+04 | 1.26E+06 | 0.00E+00 | -5.53 | -0.321 | -0.0956 | -0.417 |
| Deserts, and xeric shrublands | 4.63E+04 | 7.00E+02 | 2.00E+02 | 9.00E+02 | 4.72E+04 | -1.21E+06 | -0.207 | -0.0113 | -0.00478 | -0.0160 |
| Mediterranean forests, woodlands, and shrublands | 1.44E+06 | 2.00E+04 | 8.00E+03 | 2.80E+04 | 1.47E+06 | 2.10E+05 | -6.45 | -0.321 | -0.191 | -0.513 |
| Pasture | 7.20E+05 | 7.00E+03 | 7.00E+03 | 1.40E+04 | 7.34E+05 | -5.25E+05 | -3.22 | -0.113 | -0.167 | -0.280 |
| Temperate forests | 8.75E+06 | 1.00E+05 | 7.00E+04 | 1.70E+05 | 8.92E+06 | 7.66E+06 | -39.2 | -1.61 | -1.67 | -3.28 |
| Temperate grasslands, savannas, and shrublands | 6.18E+06 | 7.00E+04 | 5.00E+04 | 1.20E+05 | 6.30E+06 | 5.04E+06 | -27.6 | -1.13 | -1.20 | -2.32 |
| Tropical and subtropical forests | 5.15E+06 | 9.00E+04 | 1.00E+04 | 1.00E+05 | 5.25E+06 | 3.99E+06 | -23.0 | -1.45 | -0.239 | -1.69 |
| Tropical and subtropical grasslands, savannas, and shrublands | 2.93E+06 | 5.00E+04 | 7.00E+03 | 5.70E+04 | 2.99E+06 | 1.73E+06 | -13.1 | -0.804 | -0.167 | -0.971 |
| Tundra | 5.66E+06 | 4.00E+04 | 7.00E+04 | 1.10E+05 | 5.77E+06 | 4.51E+06 | -25.3 | -0.643 | -1.67 | -2.32 |
Crops are the biome with the 3rd highest welfare among the 10 I analysed. I infer increasing cropland only decreases welfare if it replaces pasture, or deserts, and xeric shrublands, which are the 2 biomes with a lower density of soil animals than crops.
Cost-effectiveness
My results imply cropland replacing tropical and subtropical grasslands, savannas, and shrublands decreases the living time of soil animals by 1.73 M animal-years per m2-year, and increases their welfare by 8.15 QALY/m2-year, of which 93.2 % comes from increasing the welfare of soil nematodes. I multiply the increase in QALY/m2-year by changes in cropland in m2-year/$ to estimate the cost-effectiveness accounting only for soil animals. I present below my calculations of the increase in cropland.
I estimated broiler welfare and cage-free reforms increase cropland by 1.98 m2-year/meat-kg and 0.113 m2-year/egg-kg. I also calculated broilers in conventional scenarios produce 15.8 meat-kg/broiler-year, and hens in cages 13.4 egg-kg/hen-year. So I deduce broiler welfare and cage-free reforms increase cropland by 31.3 m2-year/broiler-year and 1.51 m2-year/hen-year. I determined broiler welfare and cage-free corporate campaigns improve 3.00 and 10.8 chicken-year/$. So I conclude they increase cropland by 93.9 and 16.4 m2-year/$.
I stipulate buying beef increases agricultural land by 326 m2-year/meat-kg. Beef costed 2.87 $/meat-lb, 6.32 $/meat-kg, during the 1st quarter of 2025. So I estimate buying beef increases agricultural land by 51.6 m2-year/$. I assume all of this respects additional cropland (with none respecting pastures). Note one would ideally buy the beef, and then throw it into the bin. Offering it to people would tend to decrease their own consumption.
I estimate GiveWell’s top charities increase the living time of humans by 0.0157 human-year/$, which is the ratio between the period life expectancy at birth in low income countries in 2023 of 64.9 human-year/life, and the mean cost of saving a life donating to those charities in 2021 to 2023 of 4.13 k$/life. I wanted to use the mean number of lives saved per $ donated, but GiveWell does not provide data for that. I consider each human-year caused by GiveWell’s top charities increases the welfare of soil animals as much as increasing cropland by 8.70 k m2-year, which was the agricultural land per capita in low income countries in 2022. So I conclude an increase in cropland of 137 m2-year/$ increases the welfare of soil animals as cost-effectively as GiveWell’s top charities. I am underestimating the increase in cropland-years per $ due to the cohort life expectancy being longer than the period life expectancy, but overestimating it due to lives not being saved at birth, and the agricultural land per capita in low income countries having been decreasing.
I assume donating to HIPF from CEARCH increases human-years 12 times as cost-effectively as GiveWell’s top charities, as I estimate the cost-effectiveness of donating to HIPF accounting only for humans is 12 times that of GiveWell’s top charities. Multiplying that by my estimate for the increase in the living time of humans caused by GiveWell’s top charities of 0.0128 human-year/$, I infer donating to HIPF increases the living time of humans by 0.189 human-year/$. Combining this with the above increase in cropland of 8.70 k m2-year/human-year, I conclude donating to HIPF increases the welfare of soil animals as cost-effectively as increasing cropland by 1.64 k m2-year/$. I get the ratio of 12 from the mean between the lower and upper bound of 9 and 15 mentioned by Joel Tan, CEARCH’s founder and managing director, on 28 May 2025. CEARCH estimated the cost-effectiveness accounting only for effects on humans, as a fraction of that of GiveWell’s top charities, of donating to Giving What We Can (GWWC) in 2025 to be 13, that of “advocacy for top sodium control policies to control hypertension” to be 31, that of advocating for “increasing the degree to which governments respond with effective food distribution measures, continued trade, and adaptations to the agricultural sector” in “global agricultural crises [such as nuclear and volcanic winters]” to be 33 (although I estimated this should be 12.4 % as high), and that of “advocacy for sugar-sweetened beverages [SSBs] taxes to control diabetes mellitus type 2” to be 55. Joel disclaimed he thinks the cost-effectiveness estimates from CEARCH’s deep reports, such as the ones I just mentioned, as well as (similarly elaborate) estimates from other impact-focussed evaluators, are 3 times as high as they should be. This largely explains why the mean between Joel’s lower and upper bound for the marginal cost-effectiveness of HIPF is only 21.8 % (= 12/55) of CEARCH’s highest cost-effectiveness estimate respecting advocacy for taxing SSBs.
School Plates is a program aiming to increase the consumption of plant-based foods at schools and universities in the United Kingdom (UK), where the consumption of meat, fish and seafood in 2022 was 101 meat-kg/human-year. I guess 75 % of this, 75.8 meat-kg/human-year, respects lunches and dinners, which are the meals swapped to meat-free by School Plates. I assume 2 lunches/dinners per human-day, 731 per human-year. Consequently, I arrive at a consumption of meat, fish, and seafood during lunches and dinners in the UK of 0.104 meat-kg/meal. The consumption of poultry, beef and buffalo, sheep and goat, and pork in the UK in 2022 were 42.8 %, 20.8 %, 4.94 %, and 31.5 % of that of all those meats. Multiplying these fractions by the 0.104 meat-kg/meal, I conclude School Plates reduced their consumption by 0.0444, 0.0216, 0.00512, and 0.0326 meat-kg per swapped meal. They increase cropland by 12.2, 326, 370, and 17.4 m2-year/meat-kg, so I infer School Plates would decrease cropland by 10.0 m2-year per swapped meal if the animal-based foods were replaced by ones which did not require any cropland. I considered they were replaced by the same amount of food, 0.104 food-kg/meal, requiring 5.49 m2-year/food-kg, which is the mean between the values for tofu and peas, the 2 legumes analysed in the source I used to estimate the increase in cropland linked to the animal-based foods. I multiply those to determine the replacement foods require 0.569 m2-year per swapped meal, which implies School Plates decreases cropland by 9.46 m2-year per swapped meal. I calculated School Plates swapped 64.5 lunches/dinners per $ in 2023, which corresponds to decreasing cropland by 610 m2-year/$.
Veganuary is “a non-profit organisation that encourages people worldwide to try vegan for January and beyond”. I determine how much they increased cropland in 2024 roughly as I did above for School Plates. There were 17 countries with Veganuary campaigns in 2024. For simplicity, I assume the decrease in cropland per kg of consumption of animal-based foods reduced by Veganuary if these were replaced by ones which did not require any cropland matches the increase in cropland per kg of consumption of meat, fish, and seafood in the UK. I calculate Veganuary decreases cropland by 79.7 m2-year per kg of consumption of the animal-based foods they reduce if these were replaced by foods which did not require any cropland. This is based on the aforementioned assumptions for School Plates, that 1 kg of other meats increases cropland as much as 1 kg of the types of meat I mentioned above in their proportions in the consumption in the UK, and that fish and seafood do not increase cropland. Accounting for the aforementioned increase in cropland caused by the replacement foods of 5.49 m2-year/food-kg, I infer Veganuary decreases cropland by 74.2 m2-year per kg of consumption of animal-based foods they reduce. I determined Veganuary in 2024 reduced the consumption of animal-based foods by 0.336 meat-kg/$. Combining this with the decrease in cropland, I conclude Veganuary in 2024 decreased cropland by 24.9 m2-year/$.
I present below the cost-effectiveness of the above interventions accounting for target beneficiaries and soil animals. For the cost-effectiveness accounting only for humans of GiveWell's top charities, and HIPF, I assume averting 1 DALY is as good as 1 QALY, and that those charities saving a life is as good to humans as averting 51 DALYs. According to Open Philanthropy (OP), “GiveWell uses moral weights for child deaths that would be consistent with assuming 51 years of foregone life in the DALY framework (though that is not how they reach the conclusion)”. I estimate the cost-effectiveness of buying beef accounting only for beef cows assuming their welfare is 0.172 QALY/cow-year, 1/3 of RP’s mainline welfare ranges of pigs, and that their living time is 3.01 animal-day/meat-kg. I suppose the cost-effectiveness accounting only for farmed animals of School Plates in 2023, and Veganuary in 2024 were 1.20 %, and 19.4 % of that of cage-free campaigns accounting only for chickens.
| Intervention | Cost-effectiveness accounting only for target beneficiaries (QALY/$) | Cost-effectiveness accounting only for target beneficiaries as a fraction of the past cost-effectiveness of HSI | Cost-effectiveness accounting only for soil animals (QALY/$) | Cost-effectiveness accounting only for soil animals as a fraction of that accounting only for target beneficiaries | Decrease in the living time of soil animals (animal-year/$) | Cost-effectiveness accounting for target beneficiaries and soil animals (QALY/$) | Cost-effectiveness accounting for target beneficiaries and soil animals as a fraction of the past cost-effectiveness of HSI |
| Donating to HIPF | 0.148 | 0.0232% | 1.34E+04 | 9.03E+04 | 2.84E+09 | 1.34E+04 | 20.9 |
| GiveWell's top charities | 0.0123 | 0.00193% | 1.11E+03 | 9.03E+04 | 2.37E+08 | 1.11E+03 | 1.74 |
| Broiler welfare corporate campaigns | 1.67 | 0.261% | 765 | 458 | 1.62E+08 | 767 | 1.20 |
| Buying beef | 2.24E-04 | 3.50E-07 | 420 | 1.88E+06 | 8.93E+07 | 420 | 65.8% |
| Cage-free corporate campaigns | 4.59 | 0.718% | 133 | 29.0 | 2.83E+07 | 138 | 21.6% |
| Veganuary in 2024 | 0.0551 | 0.00862% | -203 | -3.69E+03 | -4.32E+07 | -203 | -31.8% |
| School Plates in 2023 | 0.890 | 0.139% | -4.98E+03 | -5.59E+03 | -1.06E+09 | -4.97E+03 | -7.79 |
I get the following cost-effectiveness accounting for target beneficiaries and soil animals as a fraction of the past cost-effectiveness of HSI:
- For donating to HIPF from CEARCH, 20.9 (13.4 kQALY/$).
- For GiveWell’s top charities, 1.74 (1.11 kQALY/$).
- For Broiler welfare corporate campaigns, 1.20 (767 QALY/$).
- For buying beef, 65.8 % (420 QALY/$).
- For cage-free corporate campaigns, 21.6 % (138 QALY/$).
- For Veganuary in 2024, -31.8 % (-203 QALY/$).
- For School Plates in 2023, -7.79 (-4.97 kQALY/$).
I am uncertain about whether each of the above is beneficial or harmful. Likewise for other interventions aiming to help vertebrates which change cropland. I estimate their effects on the target beneficiaries are negligible compared with those on soil animals, and it is unclear whether these have positive or negative lives, which results in cost-effectiveness distributions with positive and negative heavy tails. Gemini’s best guesses for the expected welfare per animal-year as a fraction of the welfare range, and my distributions for this suggest soil nematodes, mites, and springtails have negative lives with a probability of 58.7 %, 55.8 %, and 55.0 %.
I consider it very worth it to decrease the uncertainty about how interventions affect the living time of soil nematodes, mites, and springtails, and about their welfare in QALY/animal-year. In particular, it is crucial to know whether they have positive or negative lives. I am not aware of any organisations working reasonably directly on this. I wonder how much money RP or WAI would need to make some progress with targeted projects.
I suspect donating to HIPF increases welfare more cost-effectively than to the organisations working on invertebrate welfare I had recommended, although I believe these are much less likely to be harmful. I estimate donating to HIPF decreases the living time of soil nematodes, mites, and springtails by 2.84 billion animal-years per $.
I no longer believe animal welfare should be more prominently promoted by EGIs. Increasing donations to interventions whose target beneficiaries are humans is easier, resulting in more additional donations per $ spent, and I estimate the most cost-effective ones are roughly as cost-effective as HSI has been.
I am pessimistic about finding interventions which increase beef consumption much more cost-effectively than directly buying it. Companies selling beef would be underinvesting in increasing beef consumption if they could spend 1 $ to increase their revenue from it by more than 1 $ without significant changes in the revenue from other products.
I do not expect the meat-eating problem to be problematic due to positive effects on wild animals. Extending human lives, and increasing income increase the consumption of animal-based foods, and therefore the number of farmed animals with negative lives. I estimated a random person globally, and in China, India, and Nigeria in 2022 decreased the welfare of poultry birds and farmed aquatic animals 15.5, 34.6, 5.17, and 2.31 times as much as the person’s welfare. Nonetheless, I calculate GiveWell’s top charities increase the welfare of soil animals 90.3 k times as much as they increase the welfare of humans, which is a way higher ratio than the ones I just mentioned.
I would also say the Against Malaria Foundation (AMF) is beneficial. I estimated the insecticide-treated nets (ITNs) distributed by Against Malaria Foundation (AMF) in the Democratic Republic of the Congo (DRC) cause 763 times as much harm to mosquitoes as they benefit humans, which is much less than the above ratio of 90.3 k.
As I expected, my analysis indicates the impact of chicken welfare reforms is driven by effects on wild animals. I estimate broiler welfare and cage-free corporate campaigns benefit soil animals 458 and 29.0 times as much as they benefit chickens. I think the same applies to any intervention targeting vertebrates which changes the consumption of feed or food, especially if it mainly aims to increase/decrease positive/negative vertebrate-years.
My best guess is that decreasing the consumption of animal-based foods is harmful. I estimate School Plates in 2023, and Veganuary in 2024 harmed soil animals 5.59 k and 3.69 k times as much as they benefited farmed animals. Moreover, I guess arguments for decreasing the consumption of animal-based foods based on environmental considerations, and animal rights harm wild animals longterm due to encouraging wilderness preservation. In any case, I expect the nearterm effects to be the driver of the overall impact. So I would still consider decreasing the consumption of animal-based foods harmful even if it helped wild animals longterm.
Donating more and better becomes less valuable than I had suggested after accounting for effects on soil animals. The highest cost-effectiveness accounting only for effects on target beneficiaries respecting HSI’s past work is 139 (= 1/0.00718) times the 2nd highest regarding cage-free corporate campaigns. In contrast, the highest cost-effectiveness accounting for effects on target beneficiaries and soil animals respecting donating to HIPF is 12.0 (= 20.9/1.74) times the 2nd highest regarding GiveWell’s top charities. Yet, I maintain donating more and better is the best strategy to maximise impact for the vast majority of people. Donating 10 % more to an organisation 10 times as cost-effective as one one could join is 10 (= 0.1*10/0.1) times as impactful as working there if the alternative hire would be 10 % less impactful.
Effects on soil animals cannot be neglected just because they are uncertain
Here is an illustration of why I think effects on soil animals cannot be neglected just because they are uncertain. Consider an intervention aiming to decrease the consumption of animal-based foods which:
- Increases the welfare of farmed animals, the target beneficiaries, by 1 QALY with 100 % probability.
- Decreases the welfare of soil animals by 1 kQALY with 30 % probability, and by 0.001 QALY with 30 % probability.
- Increases the welfare of soil animals by 0.001 QALY with 20 % probability, and by 1 kQALY with 20 % probability.
There is lots of uncertainty about whether the effects on soil animals are very negative, barely negative, barely positive, or very positive, but I would not neglect them. They decrease welfare by 100 QALY (= 0.3*(1*10^3 + 0.001) - 0.2*(1*10^3 + 0.001)) in expectation, and therefore the intervention decreases welfare by 99.0 QALY (= 100 - 1*1) in expectation, thus being harmful.
You may well disagree with my numbers above. However, mathematically speaking, “number which is certain to be barely positive” + “number which can be very negative, barely negative, barely positive, or very positive” = “number which can be very negative, barely negative, barely positive, or very positive”, which may be very negative in expectation. I would only disregard the effects on soil animals if I considered them much smaller in expectation than those on farmed animals.
As far as I can tell, neglecting uncertain effects on soil animals despite deeming them very large in expectation may also be a reason for neglecting uncertain effects on target beneficiaries. The ratio between RP’s 5th and 95th percentile welfare range is 0.485 % (= 0.005/1.03) for pigs, 0.230 % (= 0.002/0.869) for chickens, 0.272 % (= 0.004/1.47) for octopuses, and exactly 0 for carp, bees, salmon, crayfish, shrimp, crabs, black soldier flies, and silkworms. Furthermore, these ratios underestimate the overall uncertainty due to only accounting for that in the welfare range. This is not my best guess, but I can see interventions helping farmed animals causing more harm to humans due to increasing the cost of food than they benefit farmed animals, at least if they target animals whose RP’s 5th percentile welfare range is 0.
Acknowledgements
Thanks to Anonymous Person for a comment that prompted me to take the implications of best guesses for the expected welfare of wild animals more seriously, and significantly contributed towards my decision to write this post, to Johan van den Hoogen for clarifications about van den Hoogen et al. (2019), to Michael St. Jules for feedback on the draft, to Stefan Geisen for clarifications about van den Hoogen et al. (2019), and to Yuval Rosenberg for clarifications about Rosenberg et al. (2023). I listed people’s names alphabetically. The views expressed in the post are my own.
tobycrisford 🔸 @ 2025-06-03T18:00 (+70)
Outsourcing the welfare estimates to Gemini seems like a risky move to me. It's a key part of the whole analysis, but is an extremely challenging question to begin answering. What's the reason to expect Gemini to be able to do a good job of this, given the blind spots we know current AI models still have?
I tried pasting your prompt into ChatGPT, with research mode, and the 3 value estimates it gave were all positive, rather than negative: https://chatgpt.com/share/683f34e8-8088-8006-8ba4-b719d025ac45
If I'm understanding right, this would flip all your conclusions on their head, and instead of trying to eliminate wild animal habitats, the top priority would be to increase them?
Such extreme sensitivity to highly uncertain quantities strikes me as a strong reductio ad absurdum argument against this approach to decision making on this kind of question. Otherwise we find ourselves oscillating wildly between "destroy all nature" and "destroy all humans" on the basis of each piece of new information, never being especially confident in either.
I like Bayesianism and expected value maximization as a framework for decision making under uncertainty, but when considering situations with enormous amounts of value described by extremely speculative probability estimates, I think we probably need to approach things differently (or at least adapt our priors so as to be less sensitive to these kind of problems). Something like Holden Karnofsky's approach here (which Anthony DiGiovanni shared with me on a recent post on insect suffering).
Edit: Anthony DiGiovanni doesn't actually endorse Holden Karnofsky's approach (see Anthony's comment below).
MichaelStJules @ 2025-06-03T18:35 (+23)
I'll add that even if I would make different methodological choices, I think it's still useful to highlight the scale of indirect effects on wild animals. The default in the community seems to be to ignore these effects, and there doesn't seem to be good justification for that. I think it's great that Vasco is taking these effects seriously and seeing where they might lead.
(And, as in my other comment, the conclusions and analysis could hold approximately anyway for those sufficiently pessimistic about the lives of wild invertebrates, or who give enough weight to sufficiently suffering-focused views.)
Vasco Grilo🔸 @ 2025-06-03T18:58 (+4)
Thanks, Michael!
MichaelStJules @ 2025-06-03T18:15 (+13)
I agree both with the specific point about using LLMs and the more general point about sensitivity to highly speculative and ambiguous values. I would endorse imprecise credences, and the use of approaches to decision-making with imprecise credences. See also Anthony's piece against precise Bayesianism.[1]
That being said, if you're sufficiently suffering-focused or confident that their lives are negative on average (or confident that their lives are positive on average), then you don't have to worry about this too much.
- ^
On difference-making ambiguity aversion as one natural group of approaches, see my 2024 post, my 2020 post and Greaves et al., 2022. I'm not confident these are the best approaches for dealing with imprecise credences (if averse to fanaticism).
Vasco Grilo🔸 @ 2025-06-03T18:57 (+4)
Thanks, Michael. Would the approaches you mention only recommend acting on the basis that wild animals have negative lives if the probability of this was sufficiently high? If so, why would my estimates of the probability of soil nematodes, mites, and springtails having negative lives of 58.7 %, 55.8 %, and 55.0 % be too low, but, for example, 70 % be sufficiently high?
MichaelStJules @ 2025-06-03T19:22 (+25)
It's not so much that there's a specific threshold away from 50%, it's more that if you're wildly uncertain and it's highly speculative, rather than assigning a single precise probability like 55%, you should use a range of probabilities, say 40% to 70%. This range has values on either side of 50%. Then:
- If you were difference-making ambiguity averse,[1] then both increasing their populations would look bad (possibly more bad lives in expectation) and decreasing their populations would look bad (possibly fewer good lives in expectation). You'd want to minimize these effects, by avoiding interventions with such large predictable effects on wild animal population sizes, or by hedging.
- If you were ambiguity averse (not difference-making), then I imagine you'd want to decrease their populations. The worst possibilities for animals in the near-term are those where wild invertebrates are sentient and have horrible lives in expectation and you'd want to make those less bad. But s-risks (and especially hellish existential risks) would plausibly dominate instead, if you can robustly mitigate them.
- On a different account dealing with imprecise credences, when we reduce their populations, you might say these wild animals are neither better off in expectation (in case they have good lives in expectation), nor are they worse off in expectation (in case they have bad lives in expectation), so we can ignore them, via a principle that extends the Pareto principle (Hedden, 2024).
(I'm assuming we're ruling out an average welfare of exactly 0 or assigning that negligible probability, EDIT: conditional on sentience/having any welfare at all.)
- ^
On standard accounts of difference-making ambiguity aversion, which I think are problematic. I'm less sure about the implications of other accounts. See my 2024 post.
Vasco Grilo🔸 @ 2025-06-03T20:12 (+4)
Thanks for clarifying, Michael.
(I'm assuming we're ruling out an average welfare of exactly 0 or assigning that negligible probability.)
I would agree any particular value for the welfare per animal-year has a negligible probability because my probability distribution is practically continuous, such that there is lots of values around any particular one.
tobycrisford 🔸 @ 2025-06-04T07:20 (+5)
Fascinating discussion between the two of you here, thanks.
I have one comment: I don't think their welfare being exactly 0 should have negligible probability. If we consider an animal like the soil nematode, I think there should be a significant probability assigned to the possibility that they are not sentient, unless I'm missing something?
MichaelStJules @ 2025-06-04T09:56 (+6)
Yes, absolutely right about 0 being possible and reaonably likely. Maybe I'd say "average welfare conditional on having any welfare at all". I only added that so that X% likely to be negative meant (100-X)% likely to be positive, in order to simplify the argument.
Vasco Grilo🔸 @ 2025-06-04T08:27 (+4)
Thanks, Toby! Credits go to Michael.
I think "probability of sentience"*"expected welfare conditional on sentience" >> (1 - "probability of sentience")*"expected welfare conditional on non-sentience", such that the expected welfare can be estimated from the 1st expression. However, I would say the expected welfare conditional on non-sentience is not exactly 0. For this to be the case, one would have to be certain that a welfare of exactly 0 follows from failing to satisfy the sentience criteria, which is not possible. Yet, in practice, it could still be the case that there is a decent probability mass on a welfare close to 0.
Vasco Grilo🔸 @ 2025-06-03T18:31 (+9)
Thanks for the comment, Toby.
Outsourcing the welfare estimates to Gemini seems like a risky move to me. It's a key part of the whole analysis, but is an extremely challenging question to begin answering. What's the reason to expect Gemini to be able to do a good job of this, given the blind spots we know current AI models still have?
I put little trust in Gemini's or anyone's estimates about whether soil nematodes, mites, and springtails have positive or negative lives. However, my conclusions do not depend on the specific values of Gemini's guesses. Any guesses for the hedonistic welfare per animal-year as a fraction of that of fully healthy animals which were negative, and not super close to 0 would lead to similar conclusions. In addition, my sense is that most people working on wild animal welfare would guess soil nematodes, mites, and springtails have negative lives. I have now clarified this in the post. In addition, Gemini's estimates are in close agreement with Ambitious Impact's estimate for wild bugs.
Gemini provided best guesses for soil nematodes, mites, and springtails of -67 %, -44 %, and -38 %, which are 1.60, 1.05, and 0.905 times Ambitious Impact’s estimate of -42 % for wild bugs based on their deprecated welfare points system.
If I'm understanding right, this would flip all your conclusions on their head, and instead of trying to eliminate wild animal habitats, the top priority would be to increase them?
Right.
Such extreme sensitivity to highly uncertain quantities strikes me as a strong reductio ad absurdum argument against this approach to decision making on this kind of question. Otherwise we find ourselves oscillating wildly between "destroy all nature" and "destroy all humans" on the basis of each piece of new information, never being especially confident in either.
Uncertainty about whether wild animals have positive or negative lives only directly translates into uncertainty about whether one should increase or decrease wild-animal-years at the margin, which is not absurd, and neither is my recommendation of saving human lives cost-effectively. Killing all wild animals or humans are not live options.
I like Bayesianism and expected value maximization as a framework for decision making under uncertainty, but when considering situations with enormous amounts of value described by extremely speculative probability estimates, I think we probably need to approach things differently (or at least adapt our priors so as to be less sensitive to these kind of problems). Something like Holden Karnofsky's approach here (which Anthony DiGiovanni shared with me on a recent post on insect suffering).
Approaches neglecting the effects on wild animals would be implicitly considering them negligible. For this to be the case, I think one would need an unreasonably certain prior that wild animals have welfare almost exactly equal to 0.
Anthony DiGiovanni @ 2025-06-05T09:32 (+7)
Something like Holden Karnofsky's approach here (which Anthony DiGiovanni shared with me on a recent post on insect suffering)
(Context for other readers: To be clear, I don't endorse Karnofsky's model, which I think is kind of ad hoc and doesn't address the root problem of arbitrariness in our credences. The least bad epistemic framework for addressing that problem, IMO, is imprecise probabilities (accounting for unawareness).)
tobycrisford 🔸 @ 2025-06-05T17:22 (+3)
Sorry, you did say this in the other thread as well and I should have made that clear in my comment originally. Have now edited.
MichaelStJules @ 2025-06-03T22:09 (+5)
Another potentially useful takeaway is that these interventions Vasco considered, or at least diet change interventions like Veganuary and School Plates, are not robustly positive in expectation, when considering exactly the near-term animal effects. So why would we support them?
These interventions don't seem justified by their direct cost-effectiveness, unless we have adequate reason to single out those effects and ignore or discount the effects on wild terrestrial invertebrates. We'd need a good reason to single out the direct effects, or refer to even more indirect or longer term reasons (e.g. moral circle expansion, space colonization and s-risks).
Vasco Grilo🔸 @ 2025-06-04T10:07 (+2)
Thanks, Michael.
I personally only care about the expected (posterior) impact. One can get a smaller expected impact by positing a more certain prior impact, but I do not know what would be the justification for being a priori very confident about the impact being 0.
I agree the interventions I considered are not robustly beneficial in expectation. However, I would not single out interventions changing the consumption of animal-based food (among the ones I analysed, all besides the broiler welfare and cage-free campaigns, and HSI). I estimate broiler welfare and cage-free corporate campaigns benefit soil animals 444 and 28.2 times as much as they benefit chickens.
Jim Buhler clarified what would be needed to neglect the uncertain nearterm effects of interventions targeting animals. I think the effects after 100 years or so are negligible, but that people neglecting nearterm effects due to their uncertainty should neglect more uncertain longterm effects even more.
Henry Howard🔸 @ 2025-06-04T01:29 (+25)
A Pascal's mugging by nematodes? Nematodes as utility monsters?
@tobycrisford 🔸 's points about conclusions that are extremely sensitive to small changes in highly uncertain values is very important and this post (which I don't think is parody based on the author's previous posts) is a great demonstration of the pitfalls.
I've commented before that these sorts of calculations that show astronomical but uncertain numbers for shrimp welfare or insect welfare or wild animal welfare could also lead down this nematode welfare route. It's not obvious to me why someone who concludes that shrimps are hugely morally significant would not also conclude that black soldier fly maggots are hugely morally significant, and then that nematodes are hugely morally significant.

(the organism on the far right is Giardia)
Interested to hear from Insect Welfare and Wild Animal Welfare advocates why they disagree that nematodes are the primary moral concern of planet Earth.
MichaelStJules @ 2025-06-04T04:10 (+25)
A Pascal's mugging by nematodes? Nematodes as utility monsters?
Pascal's bugging and the Rebugnant Conclusion (Sebo, 2024). :P
Interested to hear from Insect Welfare and Wild Animal Welfare advocates why they disagree that nematodes are the primary moral concern of planet Earth.
I'm sympathetic to difference-making risk aversion and difference-making ambiguity aversion (although see here) and assign nematodes a quite low probability of mattering much at all to me, low enough for now that I'm inclined to ignore them altogether (and what would have gone to nematodes instead goes to mitigating s-risks). Mites, springtails, copepods and insect larvae seem substantially more likely to matter to me, based on my beliefs about their capacities.
Still, I'd rather not go 100% on invertebrates either, also due to my difference-making sympathies. I'd deal with this like normative uncertainty and use a kind of bucket approach, like the Property Rights approach and hedging, with normative uncertainty about difference-making and approaches to dealing with uncertainty, about the nature of consciousness and moral patienthood and how to deal with it (although also see this), and about aggregation. So, roughly in practice, based on the probabilities of making a difference, probabilities of moral patienthood, attitudes towards risk and aggregation, I have a humans bucket, a mammals and birds bucket, a fish bucket, a shrimp and insects bucket, a mites, springtails and copepods bucket, and an s-risks bucket.
Vasco Grilo🔸 @ 2025-06-04T06:46 (+4)
Thanks for sharing, Michael! I would be curious to know which donations you would recommend if you fully endorsed expectational total hedonistic utilitarianism, like I do, as a moral theory (not necessarily as a decision criterion).
MichaelStJules @ 2025-06-04T10:11 (+6)
If using precise credences, then I'd be a strong longtermist (probably focusing on existential risks of some kind) or chase infinities. I haven't thought a lot from this perspective about practical donation recommendations, if I'm assuming not suffering-focused. If suffering-focused (like I actually am), then probably CLR.
Vasco Grilo🔸 @ 2025-06-04T13:36 (+3)
Thanks, Michael. For readers' reference, CLR stands for Center on Long-Term Risk.
I would say a 10^-100 chance of 10^100 QALY is as good as 1 QALY. However, even if I thought the risk of human extinction over the next 10 years was 10 % (I guess it is 10^-7), I would not conclude decreasing it would be astronomically cost-effective. One should be scope-sensitive not only to large potential benefits, but also their small probabilities. Longtermists typically come up with huge amounts of benefits (e.g. 10^50 QALY), and then independently guess a probability which is only moderately small (e.g. 10^-10), which results in huge expected benefits (e.g. 10^40 QALY). Yet, the amount of benefits is not independent from their probability. For reasonable distributions describing the benefits, I think the expected benefits coming from very large benefits will be negligible. For example, if the benefits are described by a power law distribution with tail index alpha > 0, their probability will be proportional to "benefits"^-(1 + alpha), so the expected benefits linked to a given amount of benefits will be proportional to "benefits"*"benefits"^-(1 + alpha) = "benefits"^-alpha. This decreases with benefits, so the expected benefits coming from astronomical benefits will be negligible.
Vasco Grilo🔸 @ 2025-06-04T07:59 (+9)
Hi Henry,
I personally think one should only care about expected welfare, so I would be happy to act based on a very low probability of animals' welfare being sufficiently high to matter. What is your criteria for caring about animals of a given species? Do you have a minimum probability of sentience? If so, why that specific value? RP estimated a probability of 6.8 % of adult nematodes being sentient. People routinely care about events which are much less likely, although the welfare of nematodes conditional on sentience would still have to be sufficiently high for them to matter conditional on sentience.
I confirm the post is not parody. I found that remark funny in a good way.
I would also be curious to hear from people enthusiastic about invertebrate welfare, but not nematode welfare. RP estimated a probability of 8.2 % of silkworms being sentient, which is just 1.21 (= 0.082/0.068) times their probability of adult nematodes being sentient.
Joel Tan🔸 @ 2025-06-04T05:02 (+24)
Generally, I'll say that even when I disagree with Vasco, I admire his willingness to go where the evidence/logic points to, even if the conclusion thus arrived at is extremely unwelcome (e.g. the meat eater problem for saving human lives, or the wild animal problem for the opposite).
FWIW CEARCH has previously looked into the meat eater problem and tried to quantify the downside for animal welfare (in equivalent human DALY terms) when saving a human life, while also trying to adjust for additional considerations such as wild animal suffering and bias (since we're keenly aware that self-interest/preconceived moral values pushes us to reason in a certain direction). Our conclusion is that saving human lives is net positive, but not as high as it would be if not for the meat eater problem.
I'm not sure how much I would update on Vasco's argument (convenient as it is for our GHD work), and my main uncertainties relate both to (a) neuron count (I wouldn't rely on LLMs to spit out the correct answer here, because even beyond the usual risks of hallucinations when there is a correct answer written out there, the true value for this probably doesn't even exist in the written literature); and (b) the neuron/welfare relationship (though Vasco's regression is pretty interesting, and a reasonably good first step).
Overall, I think more research in this area (and more funding for such research) is clearly merited.
Vasco Grilo🔸 @ 2025-06-04T06:28 (+4)
Thanks, Joel!
FWIW CEARCH has previously looked into the meat eater problem and tried to quantify the downside for animal welfare (in equivalent human DALY terms) when saving a human life, while also trying to adjust for additional considerations such as wild animal suffering and bias (since we're keenly aware that self-interest/preconceived moral values pushes us to reason in a certain direction). Our conclusion is that saving human lives is net positive, but not as high as it would be if not for the meat eater problem.
Have you considered sharing these calculations? Did you conclude that the impact on humans is larger than that on wild animals? I estimate the impact of donating to HIPF on wild animals is 87.6 k times that on humans.
(a) neuron count (I wouldn't rely on LLMs to spit out the correct answer here, because even beyond the usual risks of hallucinations when there is a correct answer written out there, the true value for this probably doesn't even exist in the written literature)
For my assumption that cropland replaces tropical and subtropical grasslands, savannas, and shrublands, 93.1 % of the increase in the welfare of soil animals comes from decreasing nematode-years, and Gemini's guess for the number of neurons of the modal soil nematode, a L1 juvenile caenorhabditis elegans, is 79.5 % (= 240/302) of that of an adult one established in the literature.
Overall, I think more research in this area (and more funding for such research) is clearly merited.
Agreed!
Joel Tan🔸 @ 2025-06-04T07:34 (+4)
Hi Vasco,
I think our estimates rely too much on subjective input that I don't think it would be useful to others, though I will say that the RP estimates helped eliminate one major source of uncertainty (even if too much remains).
Cheers,
Joel
Vasco Grilo🔸 @ 2025-06-04T08:55 (+4)
Hi Joel,
I wonder whether your reasons for keeping the calculations private should also make you want to keep the results private (although you only shared them in a qualitative way), as these follow from the same speculative inputs. I suggested sharing the calculations because I assume it would take little time, and could slightly update the views of a few people, and yours too if people comment on them.
Joel Tan🔸 @ 2025-06-04T09:41 (+6)
I think people who have otherwise not looked into this might reasonably update a bit on the fact that someone (us) looked into this, but fundamentally, they shouldn't update a lot or let this (or anything, really) change their minds without having looked into this themselves and satisfied their own worldview.
And to give more information on why I'd rather not publish our human-animal welfare comparisons - I try to regularly review this issue (e.g. there was a considerable revision after the RP moral weights were published, and a smaller one earlier this year), but to not touch this outside those regular revisions (I tend to let myself get sucked into spending too much time thinking about fundamental normative and epistemic issues in a way that is probably not very useful).
Publishing and inviting public suggestions/comments/criticisms would almost certainly cause me to spend too much time on this right now, in a way that would be detrimental to our other ongoing research (mainly effective giving) and our outreach/donor advisory work with non-EA donors (mainly GHD, some AW). On this issue, I'd rather just wait and see what is published this year (and your work is certainly very relevant/useful) and then re-evaluate at one go, maybe in early 2026.
Vasco Grilo🔸 @ 2025-06-04T13:24 (+4)
Thanks for clarifying, Joel! That makes a lot of sense.
Vasco Grilo🔸 @ 2025-06-04T06:07 (+13)
I consider it very worth it to decrease the uncertainty about how interventions affect the living time of soil nematodes, mites, and springtails, and about their welfare in QALY/animal-year. In particular, it is crucial to know whether they have positive or negative lives. I am not aware of any organisations working reasonably directly on this. I wonder how much money RP or WAI would need to make some progress with targeted projects.
@Bob Fischer, @Cameron Meyer Shorb 🔸, @cynthiaschuck, @Elisa Autric🔹, @mal_graham🔸, @Meghan Barrett, @William McAuliffe, and @Wladimir J. Alonso, do you have any thoughts on the above? I guess a few people would be interested in supporting such targeted projects. @KarolinaSarek🔸, I wonder whether the Animal Welfare Fund (AWF) would be open to this too. I would be happy to donate 3 k$.
MichaelStJules @ 2025-06-03T18:10 (+13)
I wonder how things would change if you had short AI timelines or otherwise assumed the impacts would be cut off after 5 or 10 years, say.
- I'd imagine HIPF would get adjusted downward a lot, if and because these are policy interventions targeting hypertension, diabetes (like you mention) and other chronic lifestyle diseases. Not only would policy changes take time to pass and be implemented, I'd guess their effects on chronic lifestyle diseases would be fairly small for several years after implementation, because those diseases take time to reverse or prevent.
- GiveWell top charities would get adjusted downwards because many of the estimated years of life saved could actually be lost to AI, or have radically different effects on wild animals. Instead of 50-70 years, we only get 5-10, say.
- Broiler welfare reforms, especially the breed component which drives the wild animal effects here, tend to have phase-in periods of multiple years, and welfare reforms in general are often estimated to last several years. We might only have a few years of effects, if any.
I estimate donating to HIPF from CEARCH is 12 times as cost-effective as GiveWell’s top charities. This is the mean between the lower and upper bound of 9 and 15 mentioned by Joel Tan, CEARCH’s founder and managing director, on 28 May 2025. Joel estimated the cost-effectiveness accounting only for effects on humans, as a fraction of that of GiveWell’s top charities, of “advocacy for top sodium control policies to control hypertension” to be 31, that of “advocacy for sugar-sweetened beverages [SSBs] taxes to control diabetes mellitus type 2” to be 55, and that of donating to Giving What We Can (GWWC) in 2025 to be 13.
Is the cost-effectiveness based on increases in life expectancies only, or also improvements in quality of life? Increases in life expectancy would be the main drivers for effects on animals, and quality of life changes would probably have minimal effects on animals.
Vasco Grilo🔸 @ 2025-06-03T22:47 (+11)
Thanks for the great points, @MichaelStJules!
I wonder how things would change if you had short AI timelines or otherwise assumed the impacts would be cut off after 5 or 10 years, say.
I like Ege Erdil's median time of 20 years until full automation of remote work, and I would not neglect impact after this. However, I feel like I should think more about this, and, for the reasons below, I would strongly update towards donating to SWP neglecting impact after 5 or 10 years.
From Vetted Causes' evaluation of SWP, "SWP also informed us that it typically takes 6 to 8 months for SWP to distribute a stunner and have it operational once an agreement has been signed". Assuming 0.583 years (= (6 + 8)/2/12) from agreements to impact, and 1 year from donations to agreements, there would be 1.58 years (= 0.583 + 1) from donations to impact. As a result, there would be 3.42 (= 5 - 1.58) and 8.42 years (= 10 - 1.58) of impact neglecting impact after 5 and 10 years. I assumed "10 years" of impact to estimate the past cost-effectiveness of HSI. Supposing the current marginal cost-effectiveness is equal to the past one including all years of impact, the current marginal cost-effectiveness of HSI neglecting impact after 5 and 10 years would be 34.2 % (= 3.42/10) and 84.2 % (= 8.42/10) of the past cost-effectiveness of HSI.
I would become much more pessimistic about my invertebrate welfare recommendations besides SWP neglecting impact after 5 or 10 years. They all involve research, so I guess they would take longer than HSI's 1.58 years from donations to impact.
I guess current donations to HIPF will support advocacy for taxing SSBs, as Joel and 5 other impact-focussed donors granted 63 k$ to that. From CEARCH's cost-effectiveness analysis (CEA) of that, "counterfactual introduction only has impact in year 5", after 4 years. The maximum benefits of the 5th, and 5th to 10th years are 0.637 % (= 1.07/168), and 3.88 % (= 6.52/168) of the total maximum benefits. As a result, neglecting impact after 5 and 10 years, I would estimate donating to HIPF to be 0.637 % and 3.88 % as cost-effective, which would be 1.86 % (= 0.00637/0.342) and 4.61 % (= 0.0388/0.842) as cost-effective as donating to HSI neglecting the impact of this after 5 and 10 years.
Is the cost-effectiveness based on increases in life expectancies only, or also improvements in quality of life? Increases in life expectancy would be the main drivers for effects on animals, and quality of life changes would probably have minimal effects on animals.
The cost-effectiveness estimates from CEARCH, and Joel's guess account for effects on mortality, morbidity, and consumption. I have now clarified the following. "I assume donating to HIPF from CEARCH increases human-years 12 times as cost-effectively as GiveWell’s top charities, as I estimate the cost-effectiveness of donating to HIPF accounting only for [mortality, morbidity, and economic effects on] humans is 12 times that of GiveWell’s top charities". I have also updated my estimate of the increase in the living time of humans caused by GiveWell's top charities. "I estimate GiveWell’s top charities increase the living time of humans by 0.0157 human-year/$, which is the ratio between the period life expectancy at birth in low income countries in 2023 of 64.9 human-year/life, and the mean cost of saving a life donating to those charities in 2021 to 2023 of 4.13 k$/life". Previously, I was assuming a period healthy life expectancy at birth of 51 years, as, according to Open Philanthropy (OP), “GiveWell uses moral weights for child deaths that would be consistent with assuming 51 years of foregone life in the DALY framework (though that is not how they reach the conclusion)”.
Michael St Jules 🔸 @ 2025-07-09T23:32 (+4)
Also, the primary beneficiaries of GiveWell-recommended charities are mostly infants and children, who eat less.
Vasco Grilo🔸 @ 2025-07-10T09:25 (+4)
Thanks, Michael. I agree that would further decrease the impact of interventions saving children if impact after 5 or 10 years dropped to 0. On the other hand, I think it is super unlikely that the agricultural land per capita in low income countries becomes negligible in 5 or 10 years even conditional on remote work being fully automated then. So I would say donating to GiveWell's top charities is still a good way of increasing agricultural land under short AI timelines. At the same time, I believe buying beef is more robust against short AI timelines.I guess the resulting increase in agricultural land happens over a few years, such that impact dropping to 0 after 5 or 10 years does not affect its cost-effectiveness.
Vasco Grilo🔸 @ 2025-07-30T07:15 (+6)
Thanks to @Michael St Jules 🔸 for noting I may be significantly underestimating the effects on soil arthropods due to only considering mites and springtails. I agree. I estimate below the annual welfare of nonarboreal ants is similar to that of soil mites and springtails. However, I estimate the annual welfare of soil nematodes is 12.8 times that of soil mites and springtails, and therefore accounting for ants would not significantly change the effects on soil nematodes, mites, and springtails. In any case, accounting for more wild animals with negative lives would reinforce my conclusion that effects on them are much larger than effects on target beneficiaries.
From Rosenberg et al. (2023), "Our estimated global population of nonarboreal ants, considered as part of the soil and plant litter habitat, is 5 × 10^16 (uncertainty range, 1 × 10^16 to 9 × 10^16) individuals". I assume the welfare per animal-year of a fully happy nonarboreal ant as a fraction of their welfare range is -38 %, as Gemini guessed for soil springtails. In addition, I suppose the welfare range of a nonarboreal ant is 0.0133 (= 1.02*0.013), 1.02 times the value in RP's post for black soldier flies (BSF) of 0.013, considering RP's estimate for the probability of sentience of ants is 1.02 (= 0.3/0.294) times their estimate for BSF, and my guess that the welfare range conditional on sentience of nonarboreal ants is similar to that of BSF. So I infer nonarboreal ants have a welfare of -0.00505 QALY/animal-year (= -0.38*0.0133), and -2.52*10^14 QALY/year (= -0.00505*5*10^16), which is -35.2 k (= -2.52*10^14/(7.16*10^9)) times my estimate for humans, 1.47 (= -2.52*10^14/(-1.72*10^14)) times my estimate for soil mites and springtails, but only 10.2 % (= -2.52*10^14/(-2.46*10^15)) of my estimate for soil nematodes, mites, and springtails.
MichaelStJules @ 2025-06-03T21:51 (+6)
If you discounted nematodes >10x or more, then SWP's HSI would come out ahead, or roughly tied with HIPF, right?
Vasco Grilo🔸 @ 2025-06-03T23:12 (+4)
Right. 93.1 % of the increase in the welfare of soil animals resulting from increasing cropland comes from decreasing nematode-years. For a welfare per nematode-year 10 % as high, the cost-effectiveness accounting for target beneficiaries and soil animals of donating to HIPF would become 3.28 times the past cost-effectiveness of HSI[1], which is 16.2 % (= 3.28/20.3) of the ratio I present in the post of 20.3.
MichaelStJules @ 2025-06-03T19:49 (+6)
For the comparison to Shrimp Welfare Project's Humane Slaughter Initiative, how long are you assuming the stunners are (counterfactually) used for? If I recall correctly, some prior estimates only assumed 1 year, which seems very conservative, and would probably make the comparisons to other opportunities here unfair.
Vasco Grilo🔸 @ 2025-06-03T20:14 (+4)
I assumed "10 years of acceleration of the adoption of electrical stunning, as used by Open Philanthropy (OP) according to Aaron Boddy, SWP’s chief operations officer".
Vasco Grilo🔸 @ 2025-06-28T19:06 (+5)
It is now unclear to me whether donating to HIPF from CEARCH is beneficial or harmful. I estimated it increases cropland due to decreasing human mortality, but I think their grants aiming to decrease the consumption of salt and sugar can easily decrease cropland due decreasing the consumption of food. I think this would be the case for taxing beef. Eating 85 g of beef is associated with losing 1 microlife, 30 min. For a causal effect 1/3 as large, one would live 10 min (= 30/3) less, which means eating 13.9 kcal (= 10/60/24*2*10^3) less for 2 k kcal/person-day. For total calorie consumption to decrease, it would suffice for one to replace beef with something with less than 16.4 kcal (= 13.9*100/85) less per 100 g, which is not much. Beef is the food requiring the most agricultural land per kcal, so decreasing its consumption tends to decrease cropland. However, sugar is among the foods requiring the least agricultural land per kcal, so it could still be the case that taxing it increases cropland due to increasing the consumption of foods requiring more agricultural land per kcal, but I do not know.
Vasco Grilo🔸 @ 2025-07-23T18:38 (+2)
I have now estimated funding HIPF increases agricultural land 78.5 % as cost-effectively as I had calculated due to increasing calorie consumption. This is a minor change, so I maintain my recommendation of funding HIPF. I also confirmed that taxing beef decreases agricultural land. I estimated that consuming unprocessed red meat in the US increases agricultural land 1.72 k times as much as it decreases it from reducing life expectancy.
Vasco Grilo🔸 @ 2025-07-26T09:25 (+4)
- I calculate soil nematodes, mites, and springtails have (in expectation) a welfare of -4.48*10^-6, -1.61*10^-5, and -2.39*10^-5 QALY/animal-year, and an annual welfare of -306 k, -14.2 k, and -10.6 k times that of humans. The annual welfare of soil nematodes being 12.8 times that of soil arthropods suggests the former have been unfairly dismissed relative to the latter by people who care a lot about wild animal welfare.
The number of neurons also suggests effects on soil nematodes are larger than those on soil arthropods. I estimate soil nematodes and arthropods have 169 and 52.3 as many neurons as humans, which implies soil nematodes have 3.23 (= 169/52.3) times as many neurons as soil arthropods.
Vasco Grilo🔸 @ 2025-06-25T17:48 (+4)
- My best guess is that decreasing the consumption of animal-based foods is harmful. I estimate School Plates in 2023, and Veganuary in 2024 harmed soil animals 5.75 k and 3.85 k times as much as they benefited farmed animals.
I forgot to account for the increase in cropland caused by the replacement foods. Thanks to @MichaelStJules for noting this! I have now updated the post assuming replacement foods require "5.49 m2-year/food-kg, which is the mean between the values for tofu and peas, the 2 legumes analysed in the source I used to estimate the increase in cropland linked to the animal-based foods". The results remained practically the same. I estimate School Plates in 2023, and Veganuary in 2024 harmed soil animals 5.42 k and 3.58 k times as much as they benefited farmed animals.
Bentham's Bulldog @ 2025-06-11T12:09 (+4)
Why do cage free reforms increase crop land?
Vasco Grilo🔸 @ 2025-06-11T17:37 (+2)
Hi Matthew,
Because they require more feed, although there is large uncertainty about whether this is the case for cage-free reforms. Below are the relevant paragraphs of the post where "I estimated broiler welfare and cage-free reforms increase cropland by 1.98 m2-year/meat-kg and 0.113 m2-year/egg-kg".
Broiler welfare reforms
From Figure 4 of Gittins et al. (2024), broilers in a conventional scenario need 2.75 feed-kg/meat-kg, and ones in a reformed (slower growth) scenario, corresponding to the European Chicken Commitment (ECC), need 3.70 feed-kg/meat-kg. So I suppose broiler welfare reforms increase feed by 0.950 feed-kg/meat-kg for no change in price.
I suppose meat from broilers in reformed scenarios costs 1.10 times as much as that from broilers in conventional scenarios, which is the mean across the Netherlands, Germany, France, Italy, Spain, and Poland according to Figure 3 of van Horne and Vissers (2022), which analysed the cost of ECC. For a price elasticity of demand for chicken meat of -0.07, as in Table 2 of Bouyssou et al. (2024), that implies broiler welfare reforms decrease the consumption of chicken meat by 1.12 %, which decreases feed by 0.0308 feed-kg/meat-kg. Subtracting this from the increase in feed for no change in price, I determine an increase in feed of 0.919 feed-kg/meat-kg.
[...]
Cage-free reforms
I asked Gemini 2.5 Pro (experimental) on 11 April 2025 about the feed conversion ratio (FCR) of current egg production. Gemini provided a range of 1.9 to 2.1 feed-kg/egg-kg for cages, and 2.0 to 2.2 feed-kg/egg-kg for barns. The means between the lower and upper bounds of the ranges for cages and barns are 2.00 and 2.10 feed-kg/egg-kg, so I assumed cage-free reforms increase feed by 0.100 feed-kg/egg-kg for no change in price. From Table 3 of Leinonen et al. (2012), which studied egg production in the United Kingdom (UK), hens in cages and barns consume 2.56 and 2.74 feed-kg/egg-kg, which imply cage-free reforms increase feed by 0.178 feed-kg/egg-kg. The lower increase suggested by Gemini’s values might be explained by the faster growth of cage-free production since 2012, which facilitates economies of scale. It makes sense hens in barns need more feed because they can move around, and therefore spend more energy.
I suppose eggs from hens in barns cost 1.10 times as much as those from hens in cages, which is the ratio between the mean cost in the 7 days before 9 April 2025 of class A eggs from hens in cages and barns in the EU of 2.60 and 2.85 €/egg-kg. For a “commonly assumed” price elasticity of demand for eggs of -0.15, that implies cage-free reforms decrease the consumption of eggs by 1.37 %, which decreases feed by 0.0274 feed-kg/egg-kg. Subtracting this from the increase in feed for no change in price, I determine an increase in feed of 0.0726 feed-kg/egg-kg.
Bentham's Bulldog @ 2025-06-11T17:41 (+4)
Why does being cage free need more feed?
Vasco Grilo🔸 @ 2025-06-11T17:57 (+3)
I guess because cage-free chickens can move around, and therefore spend more energy. Importantly, I expect the effects on soil nematodes, mites, and springtails to be larger than those on laying hens regardless of whether cage-free chickens increase or decrease cropland. I estimated cage-free reforms benefit laying hens 0.718 % as much as they benefit soil nematodes, mites, and springtails for an increase in feed of 3.63 % (= 0.0726/2.00). So, for the effects on laying hens to be larger than those on soil nematodes, mites, and springtails, the change in feed would have to be smaller than 0.0261 % (= 0.00718*0.0363) holding other factors constant, which is very small.
Vasco Grilo🔸 @ 2025-06-10T10:51 (+2)
I estimated 93.1 % of the increase in the welfare of soil animals resulting from increasing cropland comes from decreasing nematode-years. So my most important assumption is that nematodes have negative lives. I have now looked a bit more into how nematodes die, and my best guess continues to be that they have negative lives.
From Félix and Braendle (2010):
C. elegans shares its natural environment with a diverse animal community, in particular arthropods, molluscs and other nematodes, some of which also feed on microbes proliferating upon plant decomposition. Many are potential dispersal vectors as well as predators of C. elegans. Frequently co-occurring predators include fungi, which, depending on the species, invade the nematode through spores attaching to the cuticle or the intestine, or use trapping devices that immobilize the animal and perforate it (Figure 2). Nematophagous mites, springtails and nematodes are other potential predators often encountered in the C. elegans habitat.
The broader nematode community associated with C. elegans comprises microbivorous nematodes, often including other rhabditid nematodes, such as Oscheius sp., diplogastrids or panagrolaims, which likely compete with C. elegans for microbial food resources. Also found in rotting fruits are fungal-eating and predatory nematodes. Sometimes, C. elegans co-occurs with other Caenorhabditis species in the same location or even in the same few square millimetres of substrate. Given the rapid proliferation upon food availability, intraspecific competition for food is probably substantial.
From Frézal and Félix (2015):
Predators of C. elegans are also little studied. From co-occurrence in the wild and from laboratory experiments, possible natural predators include small arthropods, such as mites or springtails (Figure 2H), other nematodes, such as Pristionchus spp., and trapping nematophagous fungi (Lee and Widden, 1996; Kiontke and Sudhaus, 2006; Bento et al., 2010; Maguire et al., 2011; Félix and Duveau, 2012) (Table 1).
C. elegans also constantly interacts with a variety of obligate and non-obligate parasites, such as fungi, microsporidia, bacteria and viruses (Troemel et al., 2008; Félix et al., 2011; Félix and Duveau, 2012; Hodgkin et al., 2013) (Table 1). These parasites infect their host via the two most exposed parts of the nematode, the cuticle and the intestine. Some non-invasive bacteria form a biofilm along the nematode's cuticle or directly stick to it (Hodgkin et al., 2013). Other bacteria proliferate in the nematode gut, which may induce constipation and likely impairs nutrient uptake (Félix and Duveau, 2012). The most intrusive parasites enter and proliferate inside the nematode body. Some pierce the cuticle (e.g., Drechmeria coniospora [Couillault et al., 2004], Figure 2J), while others enter intestinal cells via the apical membrane (e.g., microsporidia and Orsay virus [Troemel et al., 2008; Félix et al., 2011]).